Overview The human visual system is extraordinary in the quantity and quality of information it supplies about the world. A glance is sufficient to describe the location, size, shape, color, and texture of objects and, if the objects are moving, their direction and speed. Equally remarkable is the fact that visual information can be discerned over a wide range of stimulus intensities, from the faint light of stars at night to bright sunlight. The next two chapters describe the molecular, cellular, and higher-order mechanisms that allow us to see. The first steps in the process of seeing are determined by the optics of the eye, the molecular mechanisms by which light energy is transduced into electrical signals in the retina, and the retinal circuitry that determines the information relayed from the eye to the lateral geniculate nucleus of the thalamus, and ultimately to the primary visual cortex in the occipital lobe. Refractive Index of a Transparent Substance. Light rays travel through air at a velocity of about 300,000 km/sec, but they travel much slower through transparent solids and liquids. The refractive index of a transparent substance is the ratio of the velocity of light in air to the velocity in the substance. The refractive index of air itself is 1.00. Thus, if light travels through a particular type of glass at a velocity of 200,000 km/sec, the refractive index of this glass is 300,000 divided by 200,000, or 1.50. Refraction of Light Rays at an Interface Between Two Media with Different Refractive Indices. When light rays traveling forward in a beam (as shown in Figure 49–1 A ) strike an interface that is perpendicular to the beam, the rays enter the second medium without deviating from their course. The only effect that occurs is decreased velocity of transmission and shorter wavelength, as shown in the figure by the shorter distances between wave fronts. If the light rays pass through an angulated interface as shown in Figure 49–1 B , the rays bend if the refractive indices of the two media are different from each other. In this particular figure, the light rays are leaving air, which has a refractive index of 1.00, and are entering a block of glass having a refractive index of 1.50.When the beam first strikes the angulated interface, the lower edge of the beam enters the glass ahead of the upper edge.The wave front in the upper portion of the beam continues to travel at a velocity of 300,000 km/sec, while that which entered the glass travels at a velocity of 200,000 km/sec. This causes the upper portion of the wave front to move ahead of the lower portion, so that the wave front is no longer vertical but angulated to the right. Because the direction in which light travels is always perpendicular to the plane of the wave front, the direction of travel of the light beam bends downward. This bending of light rays at an angulated interface is known as refraction. Note particularly that the degree of refraction increases as a function of (1) the ratio of the two refractive indices of the two transparent media and (2) the degree of angulation between the interface and the entering wave front.
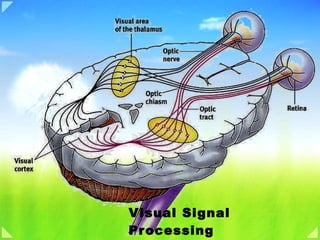
The initial stages of the mammalian visual system have the platelike organization often found in the central nervous system. The first three stages are housed in the retina; the remainder are in the brain: in the lateral geniculate bodies and the stages beyond in the cortex
Figure 26-1 Photoreceptors are located in the retina. The location of the retina within the eye is shown at left. Detail of the retina at the fovea is shown on the right (the diagram has been simplified by eliminating lateral connections mediated by interneurons; see Figure 26-6). In most of the retina light must pass through layers of nerve cells and their processes before it reaches the photoreceptors. In the center of the fovea, or foveola, these proximal neurons are shifted to the side so that light has a direct pathway to the photoreceptors. As a result, the visual image received at the foveola is the least distorted. The Retina Contains the Eye's Receptor Sheet The eye is designed to focus the visual image on the retina with minimal optical distortion. Light is focused by the cornea and the lens, then traverses the vitreous humor that fills the eye cavity before reaching photoreceptors in the retina (Figure 26-1). The retina lies in front of the pigment epithelium that lines the back of the eye. Cells in the pigment epithelium are filled with the black pigment melanin, which absorbs any light not captured by the retina. This prevents light from being reflected off the back of the eye to the retina again (which would degrade the visual image). Because the photoreceptors lie in the back of the eye, immediately in front of the pigment epithelium, all other retinal cells lie in front of the photoreceptors, closer to the lens. Therefore, light must travel through layers of other retinal neurons before striking the photoreceptors. To allow light to reach the photoreceptors without being absorbed or greatly scattered (which would distort the visual image), the axons of neurons in the proximal layers of the retina are unmyelinated so that these layers of cells are relatively transparent. Moreover, in one region of the retina, the fovea , the cell bodies of the proximal retinal neurons are shifted to the side, enabling the photoreceptors there to receive the visual image in its least distorted form (Figure 26-1). This shifting is most pronounced at the center of the fovea, the foveola. Humans therefore constantly move their eyes so that scenes of interest are projected onto the fovea. The retina also contains a region called the optic disc, where the optic nerve fibers leave the retina. This region has no photoreceptors and therefore is a blind spot in the visual field (see Figure 27-2). The projection of the visual field onto the two retinas is described in Chapter 27.
THE RETINA All this intricate superstructure exists in the interests of the retina, itself an amazing structure. It translates light into nerve signals, allows us to see under conditions that range from starlight to sunlight, discriminates wavelength so that we can see colors, and provides a precision sufficient for us to detect a human hair or speck of dust a few yards away. The retina is part of the brain, having been sequestered from it early in development but having kept its connections with the brain proper through a bundle of fibers—the optic nerve. Like many other structures in the central nervous system, the retina has the shape of a plate, in this case one about a quarter millimeter thick. It consists of three layers of nerve—cell bodies separated by two layers containing synapses made by the axons and dendrites of these cells. The tier of cells at the back of the retina contains the light receptors, the rods and cones. Rods, which are far more numerous than cones, are responsible for our vision in dim light and are out of commission in bright light. Cones do not respond to dim light but are responsible for our ability to see fine detail and for our color vision. The numbers of rods and cones vary markedly over the surface of the retina. In the very center, where our fine-detail vision is best, we have only cones. This rod-free area is called the fovea and is about half a millimeter in diameter. Cones are present throughout the retina but are most densely packed in the fovea. Because the rods and cones are at the back of the retina, the incoming light has to go through the other two layers in order to stimulate them. We do not fully understand why the retina develops in this curious backward fashion. One possible reason is the location behind the receptors of a row of cells containing a black pigment, melanin (also found in skin). Melanin mops up the light that has passed through the retina, keeping it from being reflected back and scattering around inside the eye; it has the same function as the black paint inside a camera. The melanin-containing cells also help chemically restore the light-sensitive visual pigment in the receptors after it has been bleached by light (see Chapter 8). For both functions, the melanin pigment must be close to the receptors. If the receptors were at the front of the retina, the pigment cells would have to be between them and the next layer of nerve cells, in a region already packed with axons, dendrites, and synapses. The enlarged retina at the right shows the relative positions of the three retinal layers. Surprisingly, the light has to pass through the ganglion-cell and bipolar-cell layers before it gets to the rods and cones. As it is, the layers in front of the receptors are fairly transparent and probably do not blur the image much. In the central one millimeter, however, where our vision is most acute, the consequences of even slight blurring would be disastrous, and evolution seems to have gone to some pains to alleviate it by having the other layers displaced to the side to form a ring of thicker retina, exposing the central cones so that they lie at the very front. The resulting shallow pit constitutes the fovea. Moving from back to front, we come to the middle layer of the retina, between the rods and cones and the retinal ganglion cells. This layer contains three types of nerve cells: bipolar cells, horizontal cells, and amacrine cells. Bipolar cells receive input from the receptors, as the diagram on this page shows, and many of them feed directly into the retinal ganglion cells. Horizontal cells link receptors and bipolar cells by relatively long connections that run parallel to the retinal layers; similarly, amacrine cells link bipolar cells and retinal ganglion cells. The layer of cells at the front of the retina contains the retinal ganglion cells , whose axons pass across the surface of the retina, collect in a bundle at the optic disc, and leave the eye to form the optic nerve. Each eye contains about 125 million rods and cones but only 1 million ganglion cells. In the face of this discrepancy, we need to ask how detailed visual information can be preserved. Examining the connection between cells in the retina can help resolve this problem. You can think of the information flow through the retina as following two paths: a direct path, from light receptors to bipolar cells to ganglion cells, and an indirect path, in which horizontal cells may be interposed between the receptors and bipolars, and amacrine cells between bipolars and retinal ganglion cells. (See the drawing of these direct and indirect connections on this page). These connections were already worked out in much detail by Ramon y Cajal around 1900. The direct path is highly specific or compact, in the sense that one receptor or only relatively few feed into a bipolar cell, and only one or relatively few bipolars feed into a ganglion cell. The indirect path is more diffuse, or extended, through wider lateral connections. The total area occupied by the receptors in the back layer that feed one ganglion cell in the front layer, directly and indirectly, is only about one millimeter. That area, as you may remember from Chapter 1, is the receptive field of the ganglion cell, the region of retina over which we can influence the ganglion cell's firing by light simulation. A cross section of the retina, about midway between the fovea and far periphery, where rods are more numerous than cones. From top to bottom is about one-quarter millimeter. This general plan holds for the entire retina, but the details of connections vary markedly between the fovea, which corresponds to exactly where we are looking—our center of gaze, where our ability to make out fine detail is highest—and the far outer reaches, or periphery, where vision becomes relatively crude. Between fovea and periphery, the direct part of the path from receptor to ganglion cell changes dramatically. In and near the fovea, the rule for the direct path is that a single cone feeds a single bipolar cell, and a single bipolar in turn feeds into one ganglion cell. As we go progressively farther out, however, more receptors converge on bipolars and more bipolars converge on ganglion cells. This high degree of convergence, which we find over much of the retina, together with the very compact pathway in and near the very center, helps to explain how there can be a 125:1 ratio of receptors to optic nerve fibers without our having hopelessly crude vision. The general scheme of the retinal path, with its direct and indirect components, was known for many years and its correlation with visual acuity long recognized before anyone understood the significance of the indirect path. An understanding suddenly became possible when the physiology of ganglion cells began to be studied.
Figure 11.8. Structural differences between rods and cones. Although generally similar in structure, rods (A) and cones (B) differ in their size and shape, as well as in the arrangement of the membranous disks in their outer segments. Functional Specialization of the Rod and Cone Systems Figure 11.9. The range of luminance values over which the visual system operates. At the lowest levels of illumination, only rods are activated. Cones begin to contribute to perception at about the level of starlight and are the only receptors that function under relatively bright conditions. The two types of photoreceptors, rods and cones, are distinguished by shape (from which they derive their names), the type of photopigment they contain, distribution across the retina, and pattern of synaptic connections ( Figure 11.8 ). These properties reflect the fact that the rod and cone systems (the receptors and their connections within the retina) are specialized for different aspects of vision. The rod system has very low spatial resolution but is extremely sensitive to light; it is therefore specialized for sensitivity at the expense of resolution. Conversely, the cone system has very high spatial resolution but is relatively insensitive to light; it is therefore specialized for acuity at the expense of sensitivity. The properties of the cone system also allow us to see color. The range of illumination over which the rods and cones operate is shown in Figure 11.9 . At the lowest levels of light, only the rods are activated. Such rod-mediated perception is called scotopic vision . The difficulty of making visual discriminations under very low light conditions where only the rod system is active is obvious. The problem is primarily the poor resolution of the rod system (and, to a lesser degree, the fact that there is no perception of color in dim light because the cones are not involved to a significant degree). Although cones begin to contribute to visual perception at about the level of starlight, spatial discrimination is still very poor. As illumination increases, cones become more and more dominant in determining what is seen, and they are the major determinant of perception under relatively bright conditions such as normal indoor lighting or sunlight. The contributions of rods to vision drops out nearly entirely in so-called photopic vision because their response to light saturates—that is, the membrane potential of individual rods no longer varies as a function of illumination because all of the membrane channels are closed (see Figure 11.5 ). Mesopic vision occurs in levels of light at which both rods and cones contribute—at twilight, for example. From these considerations it should be clear that most of what we think of as “seeing” is mediated by the cone system, and that loss of cone function is devastating, as occurs in elderly individuals suffering from macular degeneration ( Box C ). Individuals who have lost cone function are legally blind, whereas those who have lost rod function only experience difficulty seeing at low levels of illumination (night blindness; see Box B ). Differences in the transduction mechanisms of the two receptor types also contribute to the ability of rods and cones to respond to different ranges of light intensity. For example, rods produce a reliable response to a single photon of light, whereas more than 100 photons are required to produce a comparable response in a cone. It is not, however, that cones fail to effectively capture photons. Rather, the change in current produced by single photon capture in cones is comparatively small and difficult to distinguish from noise. Another difference is that the response of an individual cone does not saturate at high levels of steady illumination, as does the rod response. Although both rods and cones adapt to operate over a range of luminance values, the adaptation mechanisms of the cones are more effective. This difference in adaptation is apparent in the time course of the response of rods and cones to light flashes. The response of a cone, even to a bright light flash that produces the maximum change in photoreceptor current, recovers in about 200 milliseconds, more than four times faster than rod recovery. The arrangement of the circuits that transmit rod and cone information to retinal ganglion cells also contributes to the different characteristics of scotopic and photopic vision. In most parts of the retina, rod and cone signals converge on the same ganglion cells; i.e., individual ganglion cells respond to both rod and cone inputs, depending on the level of illumination. The early stages of the pathways that link rods and cones to ganglion cells, however, are largely independent. For example, the pathway from rods to ganglion cells involves a distinct class of bipolar cell (called rod bipolar) that, unlike cone bipolar cells, does not contact retinal ganglion cells. Instead, rod bipolar cells synapse with the dendritic processes of a specific class of amacrine cell that makes gap junctions and chemical synapses with the terminals of cone bipolars; these processes, in turn, make synaptic contacts on the dendrites of ganglion cells in the inner plexiform layer. Finally, the rod and cone systems differ dramatically in their degree of convergence, a factor that contributes greatly to their distinct properties. Each rod bipolar cell is contacted by a number of rods, and many rod bipolar cells contact a given amacrine cell. In contrast, the cone system is much less convergent. Thus, each retinal ganglion cell that dominates central vision (called midget ganglion cells) receives input from only one cone bipolar cell, which, in turn, is contacted by a single cone. Convergence makes the rod system a better detector of light, because small signals from many rods are pooled to generate a large response in the bipolar cell. At the same time, convergence reduces the spatial resolution of the rod system, since the source of a signal in a rod bipolar cell or retinal ganglion cell could have come from anywhere within a relatively large area of the retinal surface. The one-to-one relationship of cones to bipolar and ganglion cells is, of course, just what is required to maximize acuity. Rods Detect Dim Light Rods contain more photosensitive visual pigment than cones, enabling them to capture more light. Even more important, rods amplify light signals more than cones do. A single photon can evoke a detectable electrical response in a rod; in contrast, tens or hundreds of photons must be absorbed by a cone to evoke a similar response. In addition, the rod system is highly convergent: Many rods have synapses on the same target interneuron, known as the bipolar cell (see below). Thus, signals from the rods are pooled in the bipolar cell and reinforce one another, strengthening the signals evoked by light in individual receptors and increasing the ability of the brain to detect dim lights. In contrast, fewer cones converge on each bipolar cell. In fact, cones in the foveola have small diameters, are closely spaced, and do not converge at all; each bipolar cell receives input from a single cone.
Figure 11.10. Distribution of rods and cones in the human retina. Graph illustrates that cones are present at a low density throughout the retina, with a sharp peak in the center of the fovea. Conversely, rods are present at high density throughout most of the retina, with a sharp decline in the fovea. Boxes at top illustrate the appearance of cross sections through the outer segments of the photoreceptors at different eccentricities. The increased density of cones in the fovea is accompanied by a striking reduction in the diameter of their outer segments. Anatomical Distribution of Rods and Cones The distribution of rods and cones across the surface of the retina also has important consequences for vision ( Figure 11.10 ). Despite the fact that perception in typical daytime light levels is dominated by cone-mediated vision, the total number of rods in the human retina (91 million) far exceeds the number of cones (roughly 4.5 million). As a result, the density of rods is much greater than cones throughout most of the retina. However, this relationship changes dramatically in the fovea , a highly specialized region of the central retina that measures about 1.2 millimeters in diameter ( Figure 11.11 ). In the fovea, cone density increases almost 200-fold, reaching, at its center, the highest receptor packing density anywhere in the retina. This high density is achieved by decreasing the diameter of the cone outer segments such that foveal cones resemble rods in their appearance. The increased density of cones in the fovea is accompanied by a sharp decline in the density of rods. In fact, the central 300 µm of the fovea, called the foveola , is totally rod-free. The extremely high density of cone receptors in the fovea, and the one-to- one relationship with bipolar cells and retinal ganglion cells (see earlier), endows this region (and the cone system generally) with the capacity to mediate high visual acuity. As cone density declines with eccentricity and the degree of convergence onto retinal ganglion cells increases, acuity is markedly reduced. Just 6° eccentric to the line of sight, acuity is reduced by 75%, a fact that can be readily appreciated by trying to read the words on any line of this page beyond the word being fixated on. The restriction of highest acuity vision to such a small region of the retina is the main reason humans spend so much time moving their eyes (and heads) around—in effect directing the foveas of the two eyes to objects of interest (see Chapter 20 ). It is also the reason why disorders that affect the functioning of the fovea have such devastating effects on sight (see Box C ). Conversely, the exclusion of rods from the fovea, and their presence in high density away from the fovea, explain why the threshold for detecting a light stimulus is lower outside the region of central vision. It is easier to see a dim object (such as a faint star) by looking away from it, so that the stimulus falls on the region of the retina that is richest in rods (see Figure 11.10 ). Another anatomical feature of the fovea (which literally means “pit”) that contributes to the superior acuity of the cone system is that the layers of cell bodies and processes that overlie the photoreceptors in other areas of the retina are displaced around the fovea, and especially the foveola (see Figure 11.11 ). As a result, light rays are subjected to a minimum of scattering before they strike the photoreceptors. Finally, another potential source of optical distortion that lies in the light path to the receptors—the retinal blood vessels—are diverted away from the foveola. This central region of the fovea is therefore dependent on the underlying choroid and pigment epithelium for oxygenation and metabolic sustenance
Figure 11.6. Cyclic GMP-gated channels in the outer segment membrane are responsible for the light-induced changes in the electrical activity of photoreceptors (a rod is shown here, but the same scheme applies to cones). In the dark, cGMP levels in the outer segment are high; this molecule binds to the Na+-permeable channels in the membrane, keeping them open and allowing sodium (and other cations) to enter, thus depolarizing the cell. Exposure to light leads to a decrease in cGMP levels, a closing of the channels, and receptor hyperpolarization. Phototransduction In most sensory systems, activation of a receptor by the appropriate stimulus causes the cell membrane to depolarize, ultimately stimulating an action potential and transmitter release onto the neurons it contacts. In the retina, however, photoreceptors do not exhibit action potentials; rather, light activation causes a graded change in membrane potential and a corresponding change in the rate of transmitter release onto postsynaptic neurons. Indeed, much of the processing within the retina is mediated by graded potentials, largely because action potentials are not required to transmit information over the relatively short distances involved. Perhaps even more surprising is that shining light on a photoreceptor, either a rod or a cone, leads to membrane hyperpolarization rather than depolarization ( Figure 11.5 ). In the dark, the receptor is in a depolarized state, with a membrane potential of roughly -40 mV (including those portions of the cell that release transmitters). Progressive increases in the intensity of illumination cause the potential across the receptor membrane to become more negative, a response that saturates when the membrane potential reaches about -65 mV. Although the sign of the potential change may seem odd, the only logical requirement for subsequent visual processing is a consistent relationship between luminance changes and the rate of transmitter release from the photoreceptor terminals. As in other nerve cells, transmitter release from the synaptic terminals of the photoreceptor is dependent on voltage-sensitive Ca2+ channels in the terminal membrane. Thus, in the dark, when photoreceptors are relatively depolarized, the number of open Ca2+ channels in the synaptic terminal is high, and the rate of transmitter release is correspondingly great; in the light, when receptors are hyperpolarized, the number of open Ca2+ channels is reduced, and the rate of transmitter release is also reduced. The reason for this unusual arrangement compared to other sensory receptor cells is not known. The relatively depolarized state of photoreceptors in the dark depends on the presence of ion channels in the outer segment membrane that permit Na+ and Ca2+ ions to flow into the cell, thus reducing the degree of inside negativity ( Figure 11.6 ). The probability of these channels in the outer segment being open or closed is regulated in turn by the levels of the nucleotide cyclic guanosine monophosphate (cGMP) (as in many other second messenger systems; see Chapter 8 ). In darkness, high levels of cGMP in the outer segment keep the channels open. In the light, however, cGMP levels drop and some of the channels close, leading to hyperpolarization of the outer segment membrane, and ultimately the reduction of transmitter release at the photoreceptor synapse. The series of biochemical changes that ultimately leads to a reduction in cGMP levels begins when a photon is absorbed by the photopigment in the receptor disks. The photopigment contains a light-absorbing chromophore ( retinal , an aldehyde of vitamin A) coupled to one of several possible proteins called opsins that tune the molecule's absorption of light to a particular region of the spectrum. Indeed, it is the different protein component of the photopigment in rods and cones that contributes to the functional specialization of these two receptor types. Most of what is known about the molecular events of phototransduction has been gleaned from experiments in rods, in which the photopigment is rhodopsin ( Figure 11.7A ). When the retinal moiety in the rhodopsin molecule absorbs a photon, its configuration changes from the 11- cis isomer to all- trans retinal; this change then triggers a series of alterations in the protein component of the molecule ( Figure 11.7B ). The changes lead, in turn, to the activation of an intracellular messenger called transducin , which activates a phosphodiesterase that hydrolyzes cGMP. All of these events take place within the disk membrane. The hydrolysis by phosphodiesterase at the disk membrane lowers the concentration of cGMP throughout the outer segment, and thus reduces the number of cGMP molecules that are available for binding to the channels in the surface of the outer segment membrane, leading to channel closure. One of the important features of this complex biochemical cascade initiated by photon capture is that it provides enormous signal amplification. It has been estimated that a single light-activated rhodopsin molecule can activate 800 transducin molecules, roughly eight percent of the molecules on the disk surface. Although each transducin molecule activates only one phosphodiesterase molecule, each of these is in turn capable of catalyzing the breakdown of as many as six cGMP molecules. As a result, the absorption of a single photon by a rhodopsin molecule results in the closure of approximately 200 ion channels, or about 2% of the number of channels in each rod that are open in the dark. This number of channel closures causes a net change in the membrane potential of about 1 mV. Equally important is the fact that the magnitude of this amplification varies with the prevailing levels of illumination, a phenomenon known as light adaptation . At low levels of illumination, photoreceptors are the most sensitive to light. As levels of illumination increase, sensitivity decreases, preventing the receptors from saturating and thereby greatly extending the range of light intensities over which they operate. The concentration of Ca2+ in the outer segment appears to play a key role in the light-induced modulation of photoreceptor sensitivity. The cGMP-gated channels in the outer segment are permeable to both Na+ and Ca2+; thus, light-induced closure of these channels leads to a net decrease in the internal Ca2+ concentration. This decrease triggers a number of changes in the phototransduction cascade, all of which tend to reduce the sensitivity of the receptor to light. For example, the decrease in Ca2+ increases the activity of guanylate cyclase, the cGMP synthesizing enzyme, leading to an increase in cGMP levels. Likewise, the decrease in Ca2+ increases the affinity of the cGMP-gated channels for cGMP, reducing the impact of the light-induced reduction of cGMP levels. The regulatory effects of Ca2+ on the phototransduction cascade are only one part of the mechanism that adapts retinal sensitivity to background levels of illumination; another important contribution comes from neural interactions between horizontal cells and photoreceptor terminals. Once initiated, additional mechanisms limit the duration of this amplifying cascade and restore the various molecules to their inactivated states. The protein arrestin , for instance, blocks the ability of activated rhodopsin to activate transducin, and facilitates the breakdown of activated rhodopsin. The all- trans retinal then dissociates from the opsin, diffuses into the cytosol of the outer segment, and is transported out of the outer segment and into the pigment epithelium, where appropriate enzymes ultimately convert it to 11- cis retinal. After it is transported back into the outer segment, the 11- cis retinal recombines with opsin in the receptor disks. The recycling of rhodopsin is critically important for maintaining the light sensitivity of photoreceptors. Even under intense levels of illumination, the rate of regeneration is sufficient to maintain a significant number of active photopigment molecules.
Figure 11.7. Details of phototransduction in rod photoreceptors. (A) The molecular structure of rhodopsin, the pigment in rods. (B) The second messenger cascade of phototransduction. Light stimulation of rhodopsin in the receptor disks leads to the activation of a G-protein (transducin), which in turn activates a phosphodiesterase (PDE). The phosphodiesterase hydrolyzes cGMP, reducing its concentration in the outer segment and leading to the closure of sodium channels in the outer segment membrane. . Figure 50–6 shows movement of sodium ions in a complete electrical circuit through the inner and outer segments of the rod. The inner segment continually pumps sodium from inside the rod to the outside, thereby creating a negative potential on the inside of the entire cell. However, the outer segment of the rod, where the photoreceptor discs are located, is entirely different; here, the rod membrane, in the dark state, is very leaky to sodium ions. Therefore, positively charged sodium ions continually leak back to the inside of the rod and thereby neutralize much of the negativity on the inside of the entire cell. Thus, under normal dark conditions, when the rod is not excited, there is reduced electronegativity inside the membrane of the rod, measuring about –40 millivolts rather than the usual –70 to –80 millivolts found in most sensory receptors. Then, when the rhodopsin in the outer segment of the rod is exposed to light, the rhodopsin begins to decompose, and this decreases the outer segment membrane conductance of sodium to the interior of the rod, even though sodium ions continue to be pumped outward through the membrane of the inner segment. Thus, more sodium ions now leave the rod than leak back in. Because they are positive ions, their loss from inside the rod creates increased negativity inside the membrane, and the greater the amount of light energy striking the rod, the greater the electronegativity becomes—that is, the greater is the degree of hyperpolarization. At maximum light intensity, the membrane potential approaches –70 to –80 millivolts, which is near the equilibrium potential for potassium ions across the membrane. Duration of the Receptor Potential, and Logarithmic Relation of the Receptor Potential to Light Intensity. When a sudden pulse of light strikes the retina, the transient hyperpolarization that occurs in the rods— that is, the receptor potential that occurs—reaches a peak in about 0.3 second and lasts for more than a second. In cones, the change occurs four times as fast as in the rods. A visual image impinged on the rods of the retina for only one millionth of a second can sometimes cause the sensation of seeing the image for longer than a second. Another characteristic of the receptor potential is that it is approximately proportional to the logarithm of the light intensity. This is exceedingly important, because it allows the eye to discriminate light intensities through a range many thousand times as great as would be possible otherwise. Mechanism by Which Rhodopsin Decomposition Decreases Membrane Sodium Conductance—The Excitation “Cascade.” Under optimal conditions, a single photon of light, the smallest possible quantal unit of light energy, can cause a measurable receptor potential in a rod of about 1 millivolt. Only 30 photons of light will cause half saturation of the rod. How can such a small amount of light cause such great excitation? The answer is that the photoreceptors have an extremely sensitive chemical cascade that amplifies the stimulatory effects about a millionfold, as follows: 1. The photon activates an electron in the 11- cis retinal portion of the rhodopsin; this leads to the formation of metarhodopsin II, which is the active form of rhodopsin, as already discussed and shown in Figure 50–5. 2. The activated rhodopsin functions as an enzyme to activate many molecules of transducin, a protein present in an inactive form in the membranes of the discs and cell membrane of the rod. 3. The activated transducin activates many more molecules of phosphodiesterase. 4. Activated phosphodiesterase is another enzyme; it immediately hydrolyzes many molecules of cyclic guanosine monophosphate (cGMP), thus destroying it. Before being destroyed, the cGMP had been bound with the sodium channel protein of the rod’s outer membrane in a way that “splints” it in the open state. But in light, when phosphodiesterase hydrolyzes the cGMP, this removes the splinting and allows the sodium channels to close. Several hundred channels close for each originally activated molecule of rhodopsin. Because the sodium flux through each of these channels has been extremely rapid, flow of more than a million sodium ions is blocked by the channel closure before the channel opens again. This diminution of sodium ion flow is what excites the rod, as already discussed. 5. Within about a second, another enzyme, rhodopsin kinase, which is always present in the rod, inactivates the activated rhodopsin (the metarhodopsin II), and the entire cascade reverses back to the normal state with open sodium channels. Thus, the rods have developed an important chemical cascade that amplifies the effect of a single photon of light to cause movement of millions of sodium ions. This explains the extreme sensitivity of the rods under dark conditions. The cones are about 30 to 300 times less sensitive than the rods, but even this allows color vision at any intensity of light greater than extremely dim twilight. Photochemistry of Vision Both rods and cones contain chemicals that decompose on exposure to light and, in the process, excite the nerve fibers leading from the eye. The light-sensitive chemical in the rods is called rhodopsin; the lightsensitive chemicals in the cones, called cone pigments or color pigments, have compositions only slightly different from that of rhodopsin. In this section, we discuss principally the photochemistry of rhodopsin, but the same principles can be applied to the cone pigments. Rhodopsin-Retinal Visual Cycle, and Excitation of the Rods Rhodopsin and Its Decomposition by Light Energy. The outer segment of the rod that projects into the pigment layer of the retina has a concentration of about 40 per cent of the light-sensitive pigment called rhodopsin, or visual purple. This substance is a combination of the protein scotopsin and the carotenoid pigment retinal (also called “retinene”). Furthermore, the retinal is a particular type called 11- cis retinal. This cis form of retinal is important because only this form can bind with scotopsin to synthesize rhodopsin. When light energy is absorbed by rhodopsin, the rhodopsin begins to decompose within a very small fraction of a second, as shown at the top of Figure 50–5. The cause of this is photoactivation of electrons in the retinal portion of the rhodopsin, which leads to instantaneous change of the cis form of retinal into an all- trans form that still has the same chemical structure as the cis form but has a different physical structure— a straight molecule rather than an angulated molecule. Because the three-dimensional orientation of the reactive sites of the all- trans retinal no longer fits with the orientation of the reactive sites on the protein scotopsin, the all- trans retinal begins to pull away from the scotopsin. The immediate product is bathorhodopsin, which is a partially split combination of the all- trans retinal and scotopsin. Bathorhodopsin is extremely unstable and decays in nanoseconds to lumirhodopsin. This then decays in microseconds to metarhodopsin I, then in about a millisecond to metarhodopsin II, and finally, much more slowly (in seconds), into the completely split products scotopsin and all- trans retinal. It is the metarhodopsin II, also called activated rhodopsin, that excites electrical changes in the rods, and the rods then transmit the visual image into the central nervous system in the form of optic nerve action potential, as we discuss later. Re-formation of Rhodopsin. The first stage in re-formation of rhodopsin, as shown in Figure 50–5, is to reconvert the all- trans retinal into 11- cis retinal. This process requires metabolic energy and is catalyzed by the enzyme retinal isomerase. Once the 11- cis retinal is formed, it automatically recombines with the scotopsin to re-form rhodopsin, which then remains stable until its decomposition is again triggered by absorption of light energy. Role of Vitamin A for Formation of Rhodopsin. Note in Figure 50–5 that there is a second chemical route by which all- trans retinal can be converted into 11- cis retinal.This is by conversion of the all- trans retinal first into all- trans retinol, which is one form of vitamin A. Then the all- trans retinol is converted into 11- cis retinol under the influence of the enzyme isomerase. Finally, the 11- cis retinol is converted into 11- cis retinal, which combines with scotopsin to form new rhodopsin. Vitamin A is present both in the cytoplasm of the rods and in the pigment layer of the retina.Therefore, vitamin A is normally always available to form new retinal when needed. Conversely, when there is excess retinal in the retina, it is converted back into vitamin A, thus reducing the amount of light-sensitive pigment in the retina.We shall see later that this interconversion between retinal and vitamin A is especially important in long-term adaptation of the retina to different light intensities. Night Blindness. Night blindness occurs in any person with severe vitamin A deficiency. The simple reason for this is that without vitamin A, the amounts of retinal and rhodopsin that can be formed are severely depressed. This condition is called night blindness because the amount of light available at night is too little to permit adequate vision in vitamin A–deficient persons. For night blindness to occur, a person usually must remain on a vitamin A–deficient diet for months, because large quantities of vitamin A are normally stored in the liver and can be made available to the eyes. Once night blindness develops, it can sometimes be reversed in less than 1 hour by intravenous injection of vitamin A.
Neural Function of the Retina Neural Circuitry of the Retina Figure 50–1 shows the tremendous complexity of neural organization in the retina. To simplify this, Figure 50–11 presents the essentials of the retina’s neural connections, showing at the left the circuit in the peripheral retina and at the right the circuit in the foveal retina. The different neuronal cell types are as follows: 1. The photoreceptors themselves—the rods and cones —which transmit signals to the outer plexiform layer, where they synapse with bipolar cells and horizontal cells 2. The horizontal cells, which transmit signals horizontally in the outer plexiform layer from the rods and cones to bipolar cells 3. The bipolar cells, which transmit signals vertically from the rods, cones, and horizontal cells to the inner plexiform layer, where they synapse with ganglion cells and amacrine cells 4. The amacrine cells, which transmit signals in two directions, either directly from bipolar cells to ganglion cells or horizontally within the inner plexiform layer from axons of the bipolar cells to dendrites of the ganglion cells or to other amacrine cells 5. The ganglion cells, which transmit output signals from the retina through the optic nerve into the brain A sixth type of neuronal cell in the retina, not very prominent and not shown in the figure, is the interplexiform cell. This cell transmits signals in the retrograde direction from the inner plexiform layer to the outer plexiform layer. These signals are inhibitory and are believed to control lateral spread of visual signals by the horizontal cells in the outer plexiform layer. Their role may be to help control the degree of contrast in the visual image. The Visual Pathway from the Cones to the Ganglion Cells Functions Differently from the Rod Pathway. As is true for many of our other sensory systems, the retina has both an old type of vision based on rod vision and a new type of vision based on cone vision. The neurons and nerve fibers that conduct the visual signals for cone vision are considerably larger than those that conduct the visual signals for rod vision, and the signals are conducted to the brain two to five times as rapidly. Also, the circuitry for the two systems is slightly different, as follows. To the right in Figure 50–11 is the visual pathway from the foveal portion of the retina, representing the new, fast cone system. This shows three neurons in the direct pathway: (1) cones, (2) bipolar cells, and (3) ganglion cells. In addition, horizontal cells transmit inhibitory signals laterally in the outer plexiform layer, and amacrine cells transmit signals laterally in the inner plexiform layer. To the left in Figure 50–11 are the neural connections for the peripheral retina, where both rods and cones are present. Three bipolar cells are shown; the middle of these connects only to rods, representing the type of visual system present in many lower animals. The output from the bipolar cell passes only to amacrine cells, which relay the signals to the ganglion cells. Thus, for pure rod vision, there are four neurons in the direct visual pathway: (1) rods, (2) bipolar cells, (3) amacrine cells, and (4) ganglion cells. Also, horizontal and amacrine cells provide lateral connectivity. The other two bipolar cells shown in the peripheral retinal circuitry of Figure 50–11 connect with both rods and cones; the outputs of these bipolar cells pass both directly to ganglion cells and by way of amacrine cells. Neurotransmitters Released by Retinal Neurons. Not all the neurotransmitter chemical substances used for synaptic transmission in the retina have been entirely delineated. However, both the rods and the cones release glutamate at their synapses with the bipolar cells. Histological and pharmacological studies have shown there to be many types of amacrine cells secreting at least eight types of transmitter substances, including gamma-aminobutyric acid, glycine, dopamine, acetylcholine, and indolamine, all of which normally function as inhibitory transmitters. The transmitters of the bipolar, horizontal, and interplexiform cells are unclear, but at least some of the horizontal cells release inhibitory transmitters. Transmission of Most Signals Occurs in the Retinal Neurons by Electrotonic Conduction, Not by Action Potentials. The only retinal neurons that always transmit visual signals by means of action potentials are the ganglion cells, and they send their signals all the way to the brain through the optic nerve. Occasionally, action potentials have also been recorded in amacrine cells, although the importance of these action potentials is questionable. Otherwise, all the retinal neurons conduct their visual signals by electrotonic conduction, which can be explained as follows. Electrotonic conduction means direct flow of electric current, not action potentials, in the neuronal cytoplasm and nerve axons from the point of excitation all the way to the output synapses. Even in the rods and cones, conduction from their outer segments, where the visual signals are generated, to the synaptic bodies is by electrotonic conduction. That is, when hyperpolarization occurs in response to light in the outer segment of a rod or a cone, almost the same degree of hyperpolarization is conducted by direct electric current flow in the cytoplasm all the way to the synaptic body, and no action potential is required. Then, when the transmitter from a rod or cone stimulates a bipolar cell or horizontal cell, once again the signal is transmitted from the input to the output by direct electric current flow, not by action potentials. The importance of electrotonic conduction is that it allows graded conduction of signal strength. Thus, for the rods and cones, the strength of the hyperpolarizing output signal is directly related to the intensity of illumination; the signal is not all or none, as would be the case for each action potential. Lateral Inhibition to Enhance Visual Contrast— Function of the Horizontal Cells The horizontal cells, shown in Figure 50–11, connect laterally between the synaptic bodies of the rods and cones, as well as connecting with the dendrites of the bipolar cells. The outputs of the horizontal cells are always inhibitory. Therefore, this lateral connection provides the same phenomenon of lateral inhibition that is important in all other sensory systems—that is, helping to ensure transmission of visual patterns with proper visual contrast. This phenomenon is demonstrated in Figure 50–12, which shows a minute spot of light focused on the retina. The visual pathway from the centralmost area where the light strikes is excited, whereas an area to the side is inhibited. In other words, instead of the excitatory signal spreading widely in the retina because of spreading dendritic and axonal trees in the plexiform layers, transmission through the horizontal cells puts a stop to this by providing lateral inhibition in the surrounding areas. This is essential to allow high visual accuracy in transmitting contrast borders in the visual image. Some of the amacrine cells probably provide additional lateral inhibition and further enhancement of visual contrast in the inner plexiform layer of the retina as well.
Figure 26-7 Retinal ganglion cells respond optimally to contrast in their receptive fields. Ganglion cells have circular receptive fields, with specialized center ( pink ) and surround ( gray ) regions. On-center cells are excited when stimulated by light in the center and inhibited when stimulated in the surround; off-center cells have the opposite responses. The figure shows the responses of both types of cells to five different light stimuli (the stimulated portion of the receptive field is shown in yellow ). The pattern of action potentials fired by the ganglion cell in response to each stimulus is also shown in extracellular recordings. Duration of illumination is indicated by a bar above each record. (Adapted from Kuffler 1953.) A. On-center cells respond best when the entire central part of the receptive field is stimulated ( 3 ). These cells also respond well, but less vigorously, when only a portion of the central field is stimulated by a spot of light ( 1 ). Illumination of the surround with a spot of light ( 2 ) or ring of light ( 4 ) reduces or suppresses the cell firing, which resumes more vigorously for a short period after the light is turned off. Diffuse illumination of the entire receptive field ( 5 ) elicits a relatively weak discharge because the center and surround oppose each other's effects. B. The spontaneous firing of off-center cells is suppressed when the central area of the receptive field is illuminated ( 1, 3 ) but accelerates for a short period after the stimulus is turned off. Light shone onto the surround of the receptive field excites the cell ( 2, 4 ). THE CONCEPT OF A RECEPTIVE FIELD Narrowly defined, the term receptive field refers simply to the specific receptors that feed into a given cell in the nervous system , with one or more synapses intervening. In this narrower sense, and for vision, it thus refers simply to a region on the retina, but since Kuffler's time and because of his work the term has gradually come to be used in a far broader way. Retinal ganglion cells were historically the first example of cells whose receptive fields had a substructure: stimulating different parts of the receptive fields gave qualitatively different responses, and stimulating a large area resulted in cancellation of the effects of the subdivisions rather than addition. As presently used, receptive field tends to include a description of the substructure, or if you prefer, an account of how you have to stimulate an area to make the cell respond. When we speak of "mapping out a cell's receptive field", we often mean not simply demarcating its boundaries on the retina or the screen the animal is looking at, but also describing the substructure. As we get deeper into the central nervous system, where receptive fields tend to become more and more complex, we will find that their descriptions become increasingly elaborate. Receptive-field maps are especially useful because they allow us to predict the behavior of a cell. In the case of retinal ganglion cells, for example, suppose we stimulate an on-center cell with a long, narrow rectangle of light, just wide enough to span the receptive-field center, and long enough to go beyond the whole field, center plus surround. We would predict from the on-center map on the previous page that such a stimulus would evoke a strong response, since it covers all the center and only a small fraction of the antagonistic surround. Furthermore, from the radial symmetry of the map we can predict that the magnitude of the cell's response will be independent of the slit's orientation. Both predictions are confirmed experimentally. The receptive fields of two neighboring retinal ganglion cells will usually overlap. The smallest spot of light we can shine on the retina is likely to influence hundreds of ganglion cells, some off center and some on center. The spot will fall on the centers of some receptive fields and on the surrounds of others. My second comment concerns the important question of what a population of cells such as the output cells of the retina, are doing in response to light. To understand what ganglion cells, or any other cells in a sensory system are doing, we have to go at the problem in two ways. By mapping the receptive field, we ask how we need to stimulate to make one cell respond. But we also want to know how some particular retinal stimulus affects the entire population of ganglion cells. To answer the second question we need to begin by asking what two neighboring ganglion cells, sitting side by side in the retina, have in common. The description I have given so far of ganglion-cell receptive fields could mislead you into thinking of them as forming a mosaic of nonoverlapping little circles on the retina, like the tiles on a bathroom floor. Neighboring retinal ganglion cells in fact receive their inputs from richly overlapping and usually only slightly different arrays of receptors, as shown in the diagram on this page. This is the equivalent of saying that the receptive fields almost completely overlap You can see why by glancing at the simplified circuit in the diagram above: the cell colored purple and the one colored blue receive inputs from the overlapping regions, shown in cross section, of the appropriate colors. Because of divergence, in which one cell makes synapses with many others at each stage, one receptor can influence hundreds or thousands of ganglion cells. It will contribute to the receptive-field centers of some cells and to the surrounds of others. It will excite some cells, through their centers if they are on-center cells and through their surrounds if they are off-center cells; and it will similarly inhibit others, through their centers or surrounds. Thus a small spot shining on the retina can stir up a lot of activity, in many cells. This region was the ganglion cell's receptive field. As you might expect, the receptive field was generally centered at or very near the tip of the electrode. It soon became clear that ganglion cells were of two types, and for reasons that I will soon explain, he called them on-center cells and off-center cells . An on-center cell discharged at a markedly increased rate when a small spot was turned on anywhere within a well-defined area in or near the center of the receptive field. If you listen to the discharges of such a cell over a loudspeaker, you will first hear spontaneous firing, perhaps an occasional click, and then, when the light goes on, you will hear a barrage of impulses that sounds like a machine gun firing. We call this form of response an on response. When Kuffler moved the spot of light a small distance away from the center of the receptive field, he discovered that the light suppressed the spontaneous firing of the cell, and that when he turned off the light the cell gave a brisk burst of impulses, lasting about i second. We call this entire sequence—suppression during light and discharge following light—an off response . Exploration of the receptive field soon showed that it was cleanly subdivided into a circular on region surrounded by a much larger ring-shaped off region. The more of a given region, on or off, the stimulus filled, the greater was the response, so that maximal on responses were obtained to just the right size circular spot, and maximal off responses to a ring of just the right dimensions (inner and outer diameters). Typical recordings of responses to such stimuli are shown on this page. The center and surround regions interacted in an antagonistic way: the effect of a spot in the center was diminished by shining a second spot in the surround – as f you were telling the cell to fire faster and slower at the same time. The most impressive demonstration of this interaction between center and surround occurred when a large spot covered the entire receptive field of ganglion cell. This evoked a response that was much weaker than the response to a spot just filling the center; indeed, for some cells the effects of center to the same set stimulating the two regions cancelled each other completely. An off-center cell had just the opposite behavior. Its receptive field consisted of a small center from which off responses were obtained, and a surround that gave on responses. The two kinds of cells were intermixed and seemed to be equally common. An off-center cell discharges at its highest rate in response to a black spot on a white background, because we are now illuminating only the surround of its receptive field. In nature, dark objects are probably just as common as light ones, which may help explain why information from the retina is in the form of both on-center cells and off-center cells. If you make a spot progressively larger, the response improves until the receptive-field center is filled, then it starts to decline as more and more of the surround is included, as you can see from the graph on the next page. With a spot covering the entire field, the center either just barely wins out over the surround, or the result is a draw. This effect explains why neurophysiologists before Kuffler had such lack of success: they had recorded from these cells but had generally used diffuse light – clearly from the ideal stimulus. THE RECEPTIVE FIELDS OF RETINAL GANGLION CELLS: THE OUTPUT OF THE EYE In studying the retina we are confronted with two main problems: First, how do the rods and cones translate the light they receive into electrical, and then chemical, signals? Second, how do the subsequent cells in the next two layers—the bipolar, horizontal, amacrine, and ganglion cells—interpret this information? Before discussing the physiology of the receptors and inter-mediate cells, I want to jump ahead to describe the output of the retina—represented by the activity of the ganglion cells. The map of the receptive field of a cell is a powerful and convenient shorthand description of the cell's behavior, and thus of its output. Understanding it can help us to understand why the cells in the intermediate stages are wired up as they are, and will help explain the purpose of the direct and indirect paths. If we know what ganglion cells are telling the brain, we will have gone far toward understanding the entire retina. Around 1950, Stephen Kuffler became the first to record the responses of retinal ganglion cells to spots of light in a mammal, the cat. He was then working at the Wilmer Institute of Ophthalmology at the Johns Hopkins Hospital. In retrospect, his choice of animals was lucky because the cat's retina seems to have neither the complexity of movement responses we find in the frog or rabbit retina nor the color complications we find in the retinas of fish, birds, or monkeys. Kuffler used an optical stimulator designed by Samuel Talbot. This optical device, a modified eye doctor's ophthalmoscope, made it possible to flood the retina with steady, weak, uniform background light and also to project small, more intense stimulus spots, while directly observing both the stimulus and the electrode tip. The background light made it possible to stimulate either rods or cones or both, because only the cones work when the prevailing illumination is very bright, and only the rods work in very dim light. Kuffler recorded extracellularly from electrodes inserted through the sclera (white of the eye) directly into the retina from the front. He had little difficulty finding retinal ganglion cells, which are just under the surface and are fairly large. With a steady, diffuse background light, or even in utter darkness, most retinal ganglion cells kept up a steady, somewhat irregular firing of impulses, at rates of from 1to 2 up to about 20 impulses per second. Because one might have expected the cells to be silent in complete darkness, this firing itself came as a surprise. By searching with a small spot of light, Kuffler was able to find a region in the retina through which he could influence—increase or suppress—the retinal ganglion cell's firing. Two classes of ganglion cells can be distinguished by their responses to a small spot of light applied to the center of their receptive field (Figure 26-7). On-center ganglion cells are excited when light is directed to the center of their receptive field. Light applied to the surround inhibits the cell; the most effective inhibitory stimulus is a ring of light on the entire surround. Offcenter ganglion cells are inhibited by light applied to the center of their receptive field. However, their firing rate increases for a short period of time after the light is removed; that is, they are excited when the spot of light on the center is turned off. Light excites an offcenter ganglion cell when it is directed to the surround of the receptive field. In both types of cells the response evoked by a ring of light on the entire surround cancels the response evoked by light directed to the center almost completely. For this reason, diffuse illumination of the entire receptive field evokes only a small response in either type of cell (Figure 26-7). Not all ganglion cells have a center-surround receptive field organization. For example, a few ganglion cells respond to changes in the overall luminance of the visual field and are important in controlling pupillary reflexes (see Chapter 27). On-center and off-center ganglion cells are present in roughly equal numbers, and every photoreceptor sends output to both types. Thus, ganglion cells provide two parallel pathways for the processing of visual information. In addition, their receptive fields vary in size across the retina. In the foveal region of the primate retina, where visual acuity is greatest, the receptive fields are small, with centers that are only a few minutes of arc (60 min = 1 degree). At the periphery of the retina, where acuity is low, the fields are larger, with centers of 3°-5° (1° on the retina is equal to about 0.25 mm).
Figure 26-10 Signals from cones in the surround of a bipolar cell's receptive field are mediated by horizontal cells. Center-surround antagonism is illustrated here for an on center bipolar cell. The horizontal cell receives input from a cone in the surround of the on center bipolar cell and also has a connection with a postsynaptic cone in the center of the bipolar cell's receptive field. In the dark, horizontal cells release an inhibitory transmitter that maintains postsynaptic cones in the receptive field center in a slightly hyperpolarized state. Illumination of cones in the bipolar cell's surround hyperpolarizes those cones, which in turn hyperpolarize the postsynaptic horizontal cell. (In the dark the cones in the surround are maintained in a depolarized state and thus excite those horizontal cells.) This hyperpolarization of the horizontal cell reduces the amount of inhibitory transmitter released by the horizontal cell onto postsynaptic cones in the receptive field center, and as a result these cones become depolarized (the opposite effect of light absorption by these cones). This in turn allows the on-center bipolar cell to become hyperpolarized, the opposite effect of illumination in the receptive field center. Excitation of Some Bipolar Cells and Inhibition of Others—The Depolarizing and Hyperpolarizing Bipolar Cells Two types of bipolar cells provide opposing excitatory and inhibitory signals in the visual pathway: (1) the depolarizing bipolar cell and (2) the hyperpolarizing bipolar cell. That is, some bipolar cells depolarize when the rods and cones are excited, and others hyperpolarize. There are two possible explanations for this difference. One explanation is that the two bipolar cells are of entirely different types—one responding by depolarizing in response to the glutamate neurotransmitter released by the rods and cones, and the other responding by hyperpolarizing. The other possibility is that one of the bipolar cells receives direct excitation from the rods and cones, whereas the other receives its signal indirectly through a horizontal cell. Because the horizontal cell is an inhibitory cell, this would reverse the polarity of the electrical response. Regardless of the mechanism for the two types of bipolar responses, the importance of this phenomenon is that it allows half the bipolar cells to transmit positive signals and the other half to transmit negative signals. We shall see later that both positive and negative signals are used in transmitting visual information to the brain. Another important aspect of this reciprocal relation between depolarizing and hyperpolarizing bipolar cells is that it provides a second mechanism for lateral inhibition, in addition to the horizontal cell mechanism. Because depolarizing and hyperpolarizing bipolar cells lie immediately against each other, this provides a mechanism for separating contrast borders in the visual image, even when the border lies exactly between two adjacent photoreceptors. In contrast, the horizontal cell mechanism for lateral inhibition operates over a much greater distance. Two classes of ganglion cells can be distinguished by their responses to a small spot of light applied to the center of their receptive field (Figure 26-7). On-center ganglion cells are excited when light is directed to the center of their receptive field. Light applied to the surround inhibits the cell; the most effective inhibitory stimulus is a ring of light on the entire surround. Offcenter ganglion cells are inhibited by light applied to the center of their receptive field. However, their firing rate increases for a short period of time after the light is removed; that is, they are excited when the spot of light on the center is turned off. Light excites an off center ganglion cell when it is directed to the surround of the receptive field. In both types of cells the response evoked by a ring of light on the entire surround cancels the response evoked by light directed to the center almost completely. For this reason, diffuse illumination of the entire receptive field evokes only a small response in either type of cell (Figure 26-7). Not all ganglion cells have a center-surround receptive field organization. For example, a few ganglion cells respond to changes in the overall luminance of the visual field and are important in controlling pupillary reflexes (see Chapter 27). On-center and off-center ganglion cells are present in roughly equal numbers, and every photoreceptor sends output to both types. Thus, ganglion cells provide two parallel pathways for the processing of visual information. In addition, their receptive fields vary in size across the retina. In the foveal region of the primate retina, where visual acuity is greatest, the receptive fields are small, with centers that are only a few minutes of arc (60 min = 1 degree). At the periphery of the retina, where acuity is low, the fields are larger, with centers of 3°-5° (1° on the retina is equal to about 0.25 mm). Bipolar Cells Convey Cone Signals to Ganglion Cells Through Direct or Indirect Pathways Visual information is transferred from cones to ganglion cells along two types of pathways in the retina. Cones in the center of a ganglion cell's receptive field make direct synaptic contact with bipolar cells that in turn directly contact the ganglion cells; these connections are known as direct or vertical pathways. Signals from cones in the surround of the ganglion cell's receptive field are also conveyed to the ganglion cell through bipolar cells but only indirectly by means of horizontal and some amacrine cells; these indirect connections are called lateral pathways. Horizontal cells, which have large dendritic trees, transfer information from distant cones to bipolar cells. (Horizontal cells are also electrically coupled to each other by gap junctions and thus are able to respond to inputs from even more distant cones that contact neighboring horizontal cells.) Curiously, the horizontal cells do not appear to convey information to the bipolar cells directly, but rather by feeding back onto cones in the center of the bipolar cell's receptive field (see Figure 26-10). Some types of amacrine cells transfer information from distant bipolar cells to ganglion cells (see Figure 26-6). Most synaptic contacts in the retina are grouped in two plexiform (network-like) layers. The outer plexiform layer contains the processes of receptor, bipolar, and horizontal cells, while the inner plexiform layer contains the processes of bipolar, amacrine, and ganglion cells (see Figure 26-6). Thus the bipolar cells bridge the two plexiform layers by having processes in both. We have seen that photoreceptors respond to light with graded changes in membrane potential rather than by firing action potentials. The same is true of horizontal and bipolar cells. These cells lack voltage-gated Na+ channels capable of generating action potentials; instead they transmit signals passively (see Chapter 8). Because these cells are small and have short processes, the signals spread to their synaptic terminals without significant reduction. (Passive signal spread in cells with short processes occurs in many different parts of the brain.) In contrast, the axons of ganglion cells project considerable distances to their targets in the brain and transfer information in the form of trains of action potentials. Many types of amacrine cells also fire action potentials. Amacrine Cells and Their Functions About 30 types of amacrine cells have been identified by morphological or histochemical means. The functions of about half a dozen types of amacrine cells have been characterized, and all of them are different. One type of amacrine cell is part of the direct pathway for rod vision—that is, from rod to bipolar cells to amacrine cells to ganglion cells. Another type of amacrine cell responds strongly at the onset of a continuing visual signal, but the response dies rapidly. Other amacrine cells respond strongly at the offset of visual signals, but again, the response dies quickly. Still other amacrine cells respond when a light is turned either on or off, signaling simply a change in illumination, irrespective of direction. Still another type of amacrine cell responds to movement of a spot across the retina in a specific direction; therefore, these amacrine cells are said to be directional sensitive. In a sense, then, many or most amacrine cells are interneurons that help analyze visual signals before they ever leave the retina. Ganglion Cells and Optic Nerve Fibers Each retina contains about 100 million rods and 3 million cones; yet the number of ganglion cells is only about 1.6 million. Thus, an average of 60 rods and 2 cones converge on each ganglion cell and the optic nerve fiber leading from the ganglion cell to the brain. However, major differences exist between the peripheral retina and the central retina. As one approaches the fovea, fewer rods and cones converge on each optic fiber, and the rods and cones also become more slender. These effects progressively increase the acuity of vision in the central retina. In the center, in the central fovea, there are only slender cones—about 35,000 of them—and no rods. Also, the number of optic nerve fibers leading from this part of the retina is almost exactly equal to the number of cones, as shown to the right in Figure 50–11. This explains the high degree of visual acuity in the central retina in comparison with the much poorer acuity peripherally. Light beam Neither excited nor inhibited Excited area Inhibited area Three Types of Retinal Ganglion Cells and Their Respective Fields There are three distinct types of ganglion cells, designated W, X, and Y cells. Each of these serves a different function. Transmission of Rod Vision by the W Cells. The W cells, constituting about 40 per cent of all the ganglion cells, are small, having a diameter less than 10 micrometers, and they transmit signals in their optic nerve fibers at the slow velocity of only 8 m/sec. These ganglion cells receive most of their excitation from rods, transmitted by way of small bipolar cells and amacrine cells. They have broad fields in the peripheral retina because the dendrites of the ganglion cells spread widely in the inner plexiform layer, receiving signals from broad areas. On the basis of histology as well as physiologic experiments, the W cells seem to be especially sensitive for detecting directional movement in the field of vision, and they are probably important for much of our crude rod vision under dark conditions. Transmission of the Visual Image and Color by the X Cells. The most numerous of the ganglion cells are the X cells, representing 55 per cent of the total. They are of medium diameter, between 10 and 15 micrometers, and transmit signals in their optic nerve fibers at about 14 m/sec. The X cells have small fields because their dendrites do not spread widely in the retina. Because of this, their signals represent discrete retinal locations. Therefore, it is mainly through the X cells that the fine details of the visual image are transmitted. Also, because every X cell receives input from at least one cone, X cell transmission is probably responsible for all color vision. Function of the Y Cells to Transmit Instantaneous Changes in the Visual Image. The Y cells are the largest of all, up to 35 micrometers in diameter, and they transmit their signals to the brain at 50 m/sec or faster. They are the least numerous of all the ganglion cells, representing only 5 per cent of the total. Also, they have broad dendritic fields, so that signals are picked up by these cells from widespread retinal areas. The Y ganglion cells respond, like many of the amacrine cells, to rapid changes in the visual image— either rapid movement or rapid change in light intensity— sending bursts of signals for only small fractions of a second. These ganglion cells presumably apprise the central nervous system almost instantaneously when a new visual event occurs anywhere in the visual field, but without specifying with great accuracy the location of the event, other than to give appropriate clues that make the eyes move toward the exciting vision. Excitation of the Ganglion Cells Spontaneous, Continuous Action Potentials in the Ganglion Cells. It is from the ganglion cells that the long fibers of the optic nerve lead into the brain. Because of the distance involved, the electrotonic method of conduction employed in the rods, cones, and bipolar cells within the retina is no longer appropriate; therefore, ganglion cells transmit their signals by means of repetitive action potentials instead. Furthermore, even when unstimulated, they still transmit continuous impulses at rates varying between 5 and 40 per second. The visual signals, in turn, are superimposed onto this background ganglion cell firing. Ganglion Cells Are Specialized for the Detection of Contrasts and Rapid Changes in the Visual Image Why do ganglion cells have a center-surround receptive field organization, and why are there parallel on-center and off-center pathways? As we have just seen, ganglion cells respond only weakly to uniform illumination because of the center-surround structure of their receptive fields. They respond best when the light intensities in the center and surround are quite different. They therefore report principally the contrasts in light, rather than its absolute intensity. Most of the useful information in a visual scene is, however, contained in the pattern of contrasts. The absolute amount of light reflected by objects is relatively uninformative because it is largely determined by the intensity of the light source. Doubling the ambient light intensity will double the amount of light reflected by objects but does not alter contrasts between the objects. The center-surround organization of the receptive field of ganglion cells is therefore an adaptation for detecting useful information in the visual scene. As we shall see in Chapters 28 and 29, perception of the brightness and color of objects relies mainly on information about contrast rather than the absolute amount of light and can therefore be influenced by the contrast between an object and its surroundings. For example, the same gray ring looks much lighter against a black background than against a white one (Figure 26-8). Why does the detection of contrast start in the retina? In principle the information from photoreceptors could be sent directly to higher centers for this processing. However, signals transmitted through several relay steps to the cortex inevitably become slightly distorted. One way of minimizing the effect of transmission errors is for the retina itself to measure the difference and to transmit that information. This, in effect, is what the ganglion cell does. The firing rate of a ganglion cell provides a measure of the difference in the intensities of light illuminating the center and surround. In this way information about small differences in intensities is directly transmitted to higher centers. Parallel on-center and off-center pathways also enhance the performance of the visual system because each type of ganglion cell responds best to either rapid increases or decreases in illumination. On-center ganglion cells have a low rate of firing under dim illumination; rapid increases in firing thus signal rapid increases in light intensity in their receptive field center. In contrast, off-center ganglion cells discharge at a low rate in the light; rapid increases in firing in these cells therefore signal rapid decreases in light intensity in their receptive field center. This specialization has been demonstrated by experiments in which the function of on-center ganglion cells in awake monkeys was blocked using a pharmacological agent, aminophosphorobutyrate (APB), which selectively antagonizes transmission from photoreceptors to on-center bipolar cells. Detection of rapid increases, but not decreases, in illumination was severely impaired in these animals.
Figure 12.1. The retinal surface of the right eye, viewed with an ophthalmoscope. The optic disk is the region where the ganglion cell axons leave the retina to form the optic nerve; it is also characterized by the entrance and exit, respectively, of the ophthalmic arteries and veins that supply the retina. The macula lutea can be seen as a distinct area at the center of the optical axis (the optic disk lies nasally); the macula is the region of the retina that has the highest visual acuity. The fovea is a depression or pit about 1.5 mm in diameter that lies at the center of the macula (see Chapter 11 ). Figure 12.2. Central projections of retinal ganglion cells. Ganglion cell axons terminate in the lateral geniculate nucleus of the thalamus, the superior colliculus, the pretectum, and the hypothalamus. For clarity, only the crossing axons of the right eye are shown. Figure 12.3. The circuitry responsible for the pupillary light reflex. This pathway includes bilateral projections from the retina to the pretectum and projections from the pretectum to the Edinger-Westphal nucleus. Neurons in the Edinger-Westphal nucleus terminate in the ciliary ganglion, and neurons in the ciliary ganglion innervate the pupillary constrictor muscles. Notice that the afferent axons activate both Edinger-Westphal nuclei via the neurons in the pretectum. 12. Central Visual Pathways Overview Information supplied by the retina initiates interactions between multiple subdivisions of the brain that eventually lead to conscious perception of the visual scene, at the same time stimulating more conventional reflexes such as adjusting the size of the pupil, directing the eyes to targets of interest, and regulating homeostatic behaviors that are tied to the day/night cycle. The pathways and structures that mediate this broad range of functions are necessarily diverse. Of these, the primary visual pathway from the retina to the dorsal lateral geniculate nucleus in the thalamus and on to the primary visual cortex is the most important and certainly the most thoroughly studied component of the visual system. Different classes of neurons within this pathway encode the varieties of visual information—luminance, spectral differences, orientation, and motion—that we ultimately see. The parallel processing of different categories of visual information continues in cortical pathways that extend beyond primary visual cortex, supplying a variety of visual areas in the occipital, parietal, and temporal lobes. Visual areas in the temporal lobe are primarily involved in object recognition, whereas those in the parietal lobe are concerned with motion. Normal vision depends on the integration of information in all these cortical areas. The processes underlying visual perception are not understood and remain one of the central challenges of modern neuroscience Central Projections of Retinal Ganglion Cells Ganglion cell axons exit the retina through a circular region in its nasal part called the optic disk (or optic papilla), where they bundle together to form the optic nerve . This region of the retina contains no photoreceptors and, because it is insensitive to light, produces the perceptual phenomenon known as the blind spot ( Box A ). The optic disk is easily identified as a whitish circular area when the retina is examined with an ophthalmoscope; it also is recognized as the site from which the ophthalmic artery and veins enter (or leave) the eye ( Figure 12.1 ). In addition to being a conspicuous retinal landmark, the appearance of the optic disk is a useful gauge of intracranial pressure. The subarachnoid space surrounding the optic nerve is continuous with that of the brain; as a result, increases in intracranial pressure—a sign of serious neurological problems such as a space-occupying lesion—can be detected as a swelling of the optic disk (called papilledema). Axons in the optic nerve run a straight course to the optic chiasm at the base of the diencephalon. In humans, about 60% of these fibers cross in the chiasm, while the other 40% continue toward the thalamus and midbrain targets on the same side. Once past the chiasm, the ganglion cell axons on each side form the optic tract . Thus, the optic tract, unlike the optic nerve, contains fibers from both eyes. The partial crossing (or decussation) of ganglion cell axons at the optic chiasm allows information from corresponding points on the two retinas to be processed by approximately the same cortical site in each hemisphere, an important issue that is considered in the next section. The ganglion cell axons in the optic tract reach a number of structures in the diencephalon and midbrain ( Figure 12.2 ). The major target in the diencephalon is the dorsal lateral geniculate nucleus of the thalamus. Neurons in the lateral geniculate nucleus, like their counterparts in the thalamic relays of other sensory systems, send their axons to the cerebral cortex via the internal capsule. These axons pass through a portion of the internal capsule called the optic radiation and terminate in the primary visual (or striate ) cortex (also referred to as Brodmann's area 17 or V1 ), which lies largely along and within the calcarine fissure in the occipital lobe. The retinogeniculostriate pathway , or primary visual pathway , conveys information that is essential for most of what is thought of as seeing. Thus, damage anywhere along this route results in serious visual impairment. A second major target of the ganglion cell axons is a collection of neurons that lies between the thalamus and the midbrain in a region known as the pretectum . Although small in size compared to the lateral geniculate nucleus, the pretectum is particularly important as the coordinating center for the pupillary light reflex (i.e., the reduction in the diameter of the pupil that occurs when sufficient light falls on the retina) ( Figure 12.3 ). The initial component of the pupillary light reflex pathway is a bilateral projection from the retina to the pretectum. Pretectal neurons, in turn, project to the Edinger-Westphal nucleus , a small group of nerve cells that lies close to the nucleus of the oculomotor nerve (cranial nerve III) in the midbrain. The Edinger-Westphal nucleus contains the preganglionic parasympathetic neurons that send their axons via the oculomotor nerve to terminate on neurons in the ciliary ganglion (see Chapter 20 ). Neurons in the ciliary ganglion innervate the constrictor muscle in the iris, which decreases the diameter of the pupil when activated. Shining light in the eye thus leads to an increase in the activity of pretectal neurons, which stimulates the Edinger-Westphal neurons and the ciliary ganglion neurons they innervate, thus constricting the pupil. In addition to its normal role in regulating the amount of light that enters the eye, the pupillary reflex provides an important diagnostic tool that allows the physician to test the integrity of the visual sensory apparatus, the motor outflow to the pupillary muscles, and the central pathways that mediate the reflex. Under normal conditions, the pupils of both eyes respond identically, regardless of which eye is stimulated; that is, light in one eye produces constriction of both the stimulated eye (the direct response) and the unstimulated eye (the consensual response; see Figure 12.3 ). Comparing the response in the two eyes is often helpful in localizing a lesion. For example, a direct response in the left eye without a consensual response in the right eye suggests a problem with the visceral motor outflow to the right eye, possibly damage to the oculomotor nerve or Edinger-Westphal nucleus in the brainstem. Failure to elicit a response (either direct or indirect) to stimulation of the left eye if both eyes respond normally to stimulation of the right eye suggests damage to the sensory input from the left eye, possibly to the left retina or optic nerve. There are two other important targets of retinal ganglion cell axons. One is the suprachiasmatic nucleus of the hypothalamus, a small group of neurons at the base of the diencephalon (see Figure 28.4 ). The retinohypothalamic pathway is the route by which variation in light levels influences the broad spectrum of visceral functions that are entrained to the day/night cycle (see Chapters 21 and 28 ). The other target is the superior colliculus , a prominent structure visible on the dorsal surface of the midbrain (see Figure 1.14 ). The superior colliculus coordinates head and eye movements; its functions are considered in Chapter 20 . Figure 27-4 A simplified diagram of the projections from the retina to the visual areas of the thalamus (lateral geniculate nucleus) and midbrain (pretectum and superior colliculus). The retinal projection to the pretectal area is important for pupillary reflexes, and the projection to the superior colliculus contributes to visually guided eye movements. The projection to the lateral geniculate nucleus, and from there to the visual cortex, processes visual information for perception. The Superior Colliculus Controls Saccadic Eye Movements The superior colliculus is a structure of alternating gray cellular and white (axonal) layers lying on the roof of the midbrain. Retinal ganglion cells project directly to the superficial layers and form a map of the contralateral visual field. Cells in the superficial layers in turn project through the pulvinar nucleus of the thalamus to a broad area of the cerebral cortex, thus forming an indirect pathway from the retina to the cerebral cortex. The superior colliculus also receives extensive cortical inputs. The superficial layers receive input from the visual cortex, while deeper layers receive projections from many other areas of the cerebral cortex. These deep layers have the same map of the visual field found in the superficial layers, but the cells also respond to auditory and somatosensory stimuli as well. The locations in space represented by these multisensory inputs are aligned with one another. For example, neurons that respond to a bird flying within the contralateral visual field also will respond to its singing when it is in that same part of the field. In this way, different types of sensory information about an object are conveyed to a common region of the superior colliculus. The auditory and somatosensory inputs are adjusted to fit with the visual map in situations where the maps of these other modalities might diverge. An example of such divergence occurs when our eyes are directed to one side but our head is directed straight ahead (with respect to the body); a bird sitting where we are looking will fall in the center of the visual field but its song will locate it to one side of the auditory field. Many cells lying in the deeper layers of the colliculus also discharge vigorously before the onset of saccadic eye movements, those movements that shift the gaze rapidly from one point in the visual scene to another. These cells form a movement map in the intermediate layers of the colliculus, and this map is in register with the visual map: Cells responding to stimuli in the left visual field will discharge vigorously before a leftward-directed saccade. Although the superior colliculus receives direct retinal input, the control of these saccadic eye movements is thought to be determined more by the inputs from the cerebral cortex that reach the intermediate layers. The organization within the brain of this system for generating saccadic eye movements is considered in
Figure 27-5 The reflex pathway mediating pupillary constriction. Light signals are relayed through the midbrain pretectum, to preganglionic parasympathetic neurons in the Edinger- Westphal nucleus, and out through the parasympathetic outflow of the oculomotor nerve to the ciliary ganglion. Postganglionic neurons then innervate the smooth muscle of the pupillary sphincter.
Figure 12.10. Mixing of the pathways from the two eyes first occurs in the striate cortex. (A) Although the lateral geniculate nucleus receives inputs from both eyes, these are segregated in separate layers (see also Figure 12.14). In many species, including most primates, the inputs from the two eyes remain segregated in the ocular dominance columns of layer IV, the primary cortical target of lateral geniculate axons. Layer IV neurons send their axons to other cortical layers; it is at this stage that the information from the two eyes converges onto individual neurons. (B) Pattern of ocular dominance columns in human striate cortex. The alternating left and right eye columns in layer IV have been reconstructed from tissue sections and projected onto a photograph of the medial wall of the occipital lobe. (B from Horton and Hedley-Whyte, 1984.) Figure 27-6 The lateral geniculate nucleus is the principal subcortical site for processing visual information. Inputs from the right hemiretina of each eye project to different layers of the right lateral geniculate nucleus to create a complete representation of the left visual hemifield. Similarly, fibers from the left hemiretina of each eye project to the left lateral geniculate nucleus (not shown). The temporal crescent is not represented in contralateral inputs (see Figure 27-1). Layers 1 and 2 comprise the magnocellular layers; layers 3 through 6 comprise the parvocellular layers. All of these project to area 17, the primary visual cortex. ( C = contralateral input; I = ipsilateral input.) The Lateral Geniculate Nucleus Is the Main Terminus for Input to the Visual Cortex Ninety percent of the retinal axons terminate in the lateral geniculate nucleus, the principal subcortical structure that carries visual information to the cerebral cortex. Without this pathway visual perception is lost, although some very limited stimulus detection and movement toward objects in the visual field still is possible. This residual vision, possibly mediated by the visual pathway passing through the superior colliculus, has been called blindsight. Ganglion cells in the retina project in an orderly manner to points in the lateral geniculate nucleus, so that in each lateral geniculate nucleus there is a retinotopic representation of the contralateral half of the visual field. As in the somatosensory system, all areas of the retina are not represented equally in the nucleus. The fovea, the area of the retina with the highest density of ganglion cells, has a relatively larger representation than does the periphery of the retina. About half of the neural mass in the lateral geniculate nucleus (and in the primary visual cortex) represents the fovea and the region just around it. The much larger peripheral portions of the retina, with the lowest density of ganglion cells, are less well represented. The retinal ganglion cells in and near the centrally located fovea are densely packed to compensate for the fact that the retina's central area is less than its periphery (due to the concavity of the retina). Since this physical limitation does not exist beyond the retina, neurons in the lateral geniculate nucleus and primary visual cortex are fairly evenly distributed—connections from the more numerous neurons in the fovea are distributed over a wide area. The ratio of the area in the lateral geniculate nucleus (or in the primary visual cortex) to the area in the retina representing one degree of the visual field is called the magnification factor. In primates, including humans, the lateral geniculate nucleus contains six layers of cell bodies separated by intralaminar layers of axons and dendrites. The layers are numbered from 1 to 6, ventral to dorsal (Figure 27-6). Axons of the M and P retinal ganglion cells described in Chapter 26 remain segregated in the lateral geniculate nucleus. The two most ventral layers of the nucleus contain relatively large cells and are known as the magnocellular layers; their main retinal input is from M ganglion cells. The four dorsal layers are known as parvocellular layers and receive input from P ganglion cells. Both the magnocellular and parvocellular layers include on- and off-center cells, just as there are on- and off-center ganglion cells in the retina. An individual layer in the nucleus receives input from one eye only: fibers from the contralateral nasal hemiretina contact layers 1, 4, and 6; fibers from the ipsilateral temporal hemiretina contact layers 2, 3, and 5 (Figure 27-6). Thus, although one lateral geniculate nucleus carries complete information about the contralateral visual field, the inputs from each eye remain segregated. The inputs from the nasal hemiretina of the contralateral eye represent the complete contralateral visual hemifield, whereas the inputs from the temporal hemiretina of the ipsilateral eye represent only 90% of the hemifield because they do not include the temporal crescent (see Figure 27-1) Retinal ganglion cells have concentric receptive fields, with an antagonistic center surround organization that allows them to measure the contrast in light intensity between their receptive field center and the surround (see Chapter 26). Do the receptive fields of lateral geniculate neurons have a similar organization? David Hubel and Torsten Wiesel, who first addressed this question in the early 1960s, found that they did. They directed light onto the retina of cats and monkeys by projecting patterns of light onto a screen in front of the animal. They found that receptive fields of neurons in the lateral geniculate nucleus are similar to those in the retina: small concentric fields about one degree in diameter. As in the retina, the cells are either on-center or off-center. Like the retinal ganglion cells, cells in the lateral geniculate nucleus respond best to small spots of light in the center of their receptive field. Diffuse illumination of the whole receptive field produces only weak responses. This similarity in the receptive properties of cells in the lateral geniculate nucleus and retinal ganglion cells derives in part from the fact that each geniculate neuron receives its main retinal input from only a very few ganglion cell axons. Compared with the cerebral cortex or with many other parts of the brain, the lateral geniculates are simple structures: all or almost all of the roughly one and one half million cells in each geniculate nucleus receive input directly from optic-nerve fibers, and most (not all) of the cells send axons on to the cerebral cortex. In this sense, the lateral geniculate bodies contain only one synaptic stage, but it would be a mistake to think of them as mere relay stations. They receive fibers not only from the optic nerves but also back from the cerebral cortex, to which they project, and from the brainstem reticular formation, which plays some role in attention or arousal. Some geniculate cells with axons less than a millimeter long do not leave the geniculate but synapse locally on other geniculate cells. Despite these complicating features, single cells in the geniculate respond to light in much the same way as retinal ganglion cells, with similar on-center and off-center receptive fields and similar responses to color. In terms of visual information, then, the lateral geniculate bodies do not seem to be exerting any profound transformation, and we simply don't yet know what to make of the nonvisual inputs and the local synaptic interconnection. Magnocellular and Parvocellular Pathways Convey Different Information to the Visual Cortex We have already seen that the M ganglion cells of the retina project to the magnocellular layers of the lateral geniculate nucleus and that the P ganglion cells project to the parvocellular layers. The parvocellular and magnocellular layers in turn project to separate layers of the primary visual cortex as we shall see later in this chapter. This striking anatomical segregation has led to the view that these separate sequences of retinal ganglion, lateral geniculate, and visual cortical cells can be regarded as two parallel pathways, referred to as the M and P pathways. As indicated in Table 27-1, there are striking differences between cells in the M and P pathways. The most prominent difference between the cells in the lateral geniculate nucleus is their sensitivity to color contrast. The P cells respond to changes in color (red/green and blue/ yellow) regardless of the relative brightness of the colors, whereas M cells respond weakly to changes of color when the brightness of the color is matched. Luminance contrast is a measure of the difference between the brightest and darkest parts of the stimulus—M cells respond when contrast is as low as 2%, whereas P cells rarely respond to contrasts less than 10%. The M and P cells also differ in their response to spatial and temporal frequency. Spatial frequency is the number of repetitions of a pattern over a given distance. For example, alternating light and dark bars each occurring 10 times over a visual angle of one degree have a spatial frequency of 10 cycles per degree. Temporal frequency is how rapidly the pattern changes over time; turning the bars of a grating on and off 10 times per second would produce a temporal frequency of 10 Hz. The M cells tend to have lower spatial resolution and higher temporal resolution than P cells. One way to explore further the contribution of the M and P pathways is by selectively removing one or the other in a monkey and then measuring the monkey's ability to perform a task that is thought to depend on the ablated pathway. Because the M and P cells are in different layers in the lateral geniculate nucleus, removal of a pathway is possible through localized chemical lesions (Figure 27-7).
Figure 27-9 Each half of the visual field is represented in the contralateral primary visual cortex. In humans the primary visual cortex is located at the posterior pole of the cerebral hemisphere and lies almost exclusively on the medial surface. (In some individuals it is shifted so that part of it extends onto the lateral surface.) Areas in the primary visual cortex are devoted to specific parts of the visual field, as indicated by the corresponding numbers. The upper fields are mapped below the calcarine fissure, and the lower fields above it. The striking aspect of this map is that about half of the neural mass is devoted to representation of the fovea and the region just around it. This area has the greatest visual acuity. The Primary Visual Cortex Organizes Simple Retinal Inputs Into the Building Blocks of Visual Images The first point in the visual pathway where the receptive fields of cells are significantly different from those of cells in the retina is the primary visual cortex, also called visual area 1 (abbreviated V1). This region of cortex, Brodmann's area 17, is also called the striate cortex because it contains a prominent stripe of white matter in layer 4, the stria of Gennari , consisting of myelinated axons. Like the lateral geniculate nucleus and superior colliculus, the primary visual cortex in each cerebral hemisphere receives information exclusively from the contralateral half of the visual field (Figure 27-9). Organization and Function of the Visual Cortex Figures 51–2 and 51–3 show the visual cortex located primarily on the medial aspect of the occipital lobes. Like the cortical representations of the other sensory systems, the visual cortex is divided into a primary visual cortex and secondary visual areas. Primary Visual Cortex. The primary visual cortex (see Figure 51–2) lies in the calcarine fissure area, extending forward from the occipital pole on the medial aspect of each occipital cortex. This area is the terminus of direct visual signals from the eyes. Signals from the macular area of the retina terminate near the occipital pole, as shown in Figure 51–2, while signals from the more peripheral retina terminate at or in concentric half circles anterior to the pole but still along the calcarine fissure on the medial occipital lobe. The upper portion of the retina is represented superiorly and the lower portion inferiorly. Note in the figure the especially large area that represents the macula. It is to this region that the retinal fovea transmits its signals. The fovea is responsible for the highest degree of visual acuity. Based on retinal area, the fovea has several hundred times as much representation in the primary visual cortex as do the most peripheral portions of the retina. The primary visual cortex is also called visual area I . Still another name is the striate cortex because this area has a grossly striated appearance. Secondary Visual Areas of the Cortex. The secondary visual areas, also called visual association areas, lie lateral, anterior, superior, and inferior to the primary visual cortex. Most of these areas also fold outward over the lateral surfaces of the occipital and parietal cortex, as shown in Figure 51–3. Secondary signals are transmitted to these areas for analysis of visual meanings. For instance, on all sides of the primary visual cortex is Brodmann’s area 18 (see Figure 51–3), which is where virtually all signals from the primary visual cortex pass next. Therefore, Brodmann’s area 18 is called visual area II, or simply V-2.The other, more distant secondary visual areas have specific designations—V-3, V-4, and so forth—up to more than a dozen areas. The importance of all these areas is that various aspects of the visual image are progressively dissected and analyzed. Two Major Pathways for Analysis of Visual Information— (1) The Fast “Position” and “Motion” Pathway; (2) The Accurate Color Pathway Figure 51–3 shows that after leaving the primary visual cortex, the visual information is analyzed in two major pathways in the secondary visual areas. 1. Analysis of Third-Dimensional Position, Gross Form, and Motion of Objects. One of the analytical pathways, demonstrated in Figure 51–3 by the black arrows, analyzes the third-dimensional positions of visual objects in the space around the body. This pathway also analyzes the gross physical form of the visual scene as well as motion in the scene. In other words, this pathway tells where every object is during each instant and whether it is moving. After leaving the primary visual cortex, the signals flow generally into the posterior midtemporal area and upward into the broad occipitoparietal cortex. At the anterior border of the parietal cortex, the signals overlap with signals from the posterior somatic association areas that analyze threedimensional aspects of somatosensory signals. The signals transmitted in this position-form-motion pathway are mainly from the large Y optic nerve fibers of the retinal Y ganglion cells, transmitting rapid signals but depicting only black and white with no color. 2. Analysis of Visual Detail and Color. The red arrows in Figure 51–3, passing from the primary visual cortex into secondary visual areas of the inferior, ventral, and medial regions of the occipital and temporal cortex, show the principal pathway for analysis of visual detail. Separate portions of this pathway specifically dissect out color as well. Therefore, this pathway is concerned with such visual feats as recognizing letters, reading, determining the texture of surfaces, determining detailed colors of objects, and deciphering from all this information what the object is and what it means.
Figure 12.15. Subdivisions of the extrastriate cortex in the macaque monkey. (A) Each of the subdivisions indicated in color contains neurons that respond to visual stimulation. Many are buried in sulci, and the overlying cortex must be removed in order to expose them. Some of the more extensively studied extrastriate areas are specifically identified (V2, V3, V4, and MT). V1 is the primary visual cortex; MT is the middle temporal area. (B) The arrangement of extrastriate and other areas of neocortex in a flattened view of the monkey neocortex. There are at least 25 areas that are predominantly or exclusively visual in function, plus 7 other areas suspected to play a role in visual processing. (A after Maunsell and Newsome, 1987; B after Felleman and Van Essen, 1991.) The Functional Organization of Extrastriate Visual Areas Anatomical and electrophysiological studies in monkeys have led to the discovery of a multitude of areas in the occipital, parietal, and temporal lobes that are involved in processing visual information ( Figure 12.15 ). Each of these areas contains a map of visual space, and each is largely dependent on the primary visual cortex for its activation. The response properties of the neurons in some of these areas suggest that they are specialized for different aspects of the visual scene. For example, the middle temporal area ( MT ) contains neurons that respond selectively to the direction of a moving edge without regard to its color. In contrast, neurons in another cortical area called V4 respond selectively to the color of a visual stimulus without regard to its direction of movement. These physiological findings are supported by behavioral evidence; thus, damage to area MT leads to a specific impairment in a monkey's ability to perceive the direction of motion in a stimulus pattern, while other aspects of visual perception remain intact. Recent functional imaging studies have indicated a similar arrangement of visual areas within human extrastriate cortex. Using retinotopically restricted stimuli, it has been possible to localize at least 10 separate representations of the visual field ( Figure 12.16 ). One of these areas exhibits a large motion-selective signal, suggesting that it is the homologue of the motion-selective middle temporal area described in monkeys. Another area exhibits color-selective responses, suggesting that it may be similar to V4 in nonhuman primates. A role for these areas in the perception of motion and color, respectively, is further supported by evidence for increases in activity not only during the presentation of the relevant stimulus, but also during periods when subjects experience motion or color afterimages. The clinical description of selective visual deficits after localized damage to various regions of extrastriate cortex also supports functional specialization of extrastriate visual areas in humans. For example, a well-studied patient who suffered a stroke that damaged the extrastriate region thought to be comparable to area MT in the monkey was unable to appreciate the motion of objects. The neurologist who treated her noted that she had difficulty in pouring tea into a cup because the fluid seemed to be “frozen.” In addition, she could not stop pouring at the right time because she was unable to perceive when the fluid level had moved to the brim. The patient also had trouble following a dialogue because she could not follow the movements of the speaker's mouth. Crossing the street was potentially terrifying because she couldn't judge the movement of approaching cars. As the patient related, “When I'm looking at the car first, it seems far away. But then, when I want to cross the road, suddenly the car is very near.” Her ability to perceive other features of the visual scene, such as color and form, was intact. Another example of a specific visual deficit as a result of damage to extrastriate cortex is cerebral achromatopsia . These patients lose the ability to see the world in color, whereas other aspects of vision remain in good working order. The normal colors of a visual scene are described as being replaced by “dirty” shades of gray, much like looking at a poor quality black-and-white movie. Achromatopsic individuals know the normal colors of objects—that a school bus is yellow, an apple red—but can no longer see them. Thus, when asked to draw objects from memory, they have no difficulty with shapes but are unable to appropriately color the objects they have represented. It is important to distinguish this condition from the color blindness that arises from the congenital absence of one or more cone pigments in the retina (see Chapter 11 ). In achromatopsia, the three types of cones are functioning normally; it is damage to specific extrastriate cortical areas that renders the patient unable to use the information supplied by the retina. Based on the anatomical connections between visual areas, differences in electrophysiological response properties, and the effects of cortical lesions, a consensus has emerged that extrastriate cortical areas are organized into two largely separate systems that eventually feed information into cortical association areas in the temporal and parietal lobes (see Chapter 26 ). One system, called the ventral stream, includes area V4 and leads from the striate cortex into the inferior part of the temporal lobe. This system is thought to be responsible for high-resolution form vision and object recognition. The dorsal stream, which includes the middle temporal area, leads from striate cortex into the parietal lobe. This system is thought to be responsible for spatial aspects of vision, such as the analysis of motion, and positional relationships between objects in the visual scene ( Figure 12.17 ). The functional dichotomy between these two streams is supported by observations on the response properties of neurons and the effects of selective cortical lesions. Neurons in the ventral stream exhibit properties that are important for object recognition, such as selectivity for shape, color, and texture. At the highest levels in this pathway, neurons exhibit even greater selectivity, responding preferentially to faces and objects (see Chapter 26 ). In contrast, those in the dorsal stream are not tuned to these properties, but show selectivity for direction and speed of movement. Consistent with this interpretation, lesions of the parietal cortex severely impair an animal's ability to distinguish objects on the basis of their position, while having little effect on its ability to perform object recognition tasks. In contrast, lesions of the inferotemporal cortex produce profound impairments in the ability to perform recognition tasks but no impairment in spatial tasks. These effects are remarkably similar to the syndromes associated with damage to the parietal and temporal lobe in humans (see Chapters 26 and 27 ). What, then, is the relationship between these “higher order” extrastriate visual pathways and the magno- and parvocellular pathways that supply the primary visual cortex? Not long ago, it seemed that these intracortical pathways were simply a continuation of the geniculostriate pathways—that is, the magnocellular pathway provided input to the dorsal stream and the parvocellular pathway provided input to the ventral stream. However, more recent work has indicated that the situation is more complicated. The temporal pathway clearly has access to the information conveyed by both the magno- and parvocellular streams; and the parietal pathway, while dominated by inputs from the magnocellular stream, also receives inputs from the parvocellular stream. Thus, interaction and cooperation between the magno- and parvocellular streams appear to be the rule in complex visual perceptions.
Figure 12.16. Localization of multiple visual areas in the human brain using f MRI. (A,B) Lateral and medial views (respectively) of the human brain, illustrating the location of primary visual cortex (V1) and additional visual areas V2, V3, VP (ventral posterior area), V4, MT (middle temporal area), and MST (medial superior temporal area). (C) Unfolded and flattened view of retinotopically defined visual areas in the occipital lobe. Dark grey areas correspond to cortical regions that were embedded in sulci; light regions correspond to regions that were located on the surface of gyri. Visual areas in humans show a close resemblance to visual areas originally defined in monkeys (compare with Figure 12.15 ). (After Sereno et al., 1995.)
Figure 27-10 The primary visual cortex has distinct anatomical layers, each with characteristic synaptic connections. (Adapted from Lund 1988.) A. Most afferent fibers from the lateral geniculate nucleus terminate in layer 4. The axons of cells in the parvocellular layers ( P ) terminate primarily in layer 4Cβ, with minor inputs to 4A and 1, while the axons of cells in the magnocellular layers ( M ) terminate primarily in layer 4Cα. Collaterals of both types of cells also terminate in layer 6. Cells of the intralaminar regions ( I ) of the lateral geniculate nucleus terminate in the blob regions of layers 2 and 3. B. Several types of neurons make up the primary visual cortex. Spiny stellate and pyramidal cells, both of which have spiny dendrites, are excitatory. Smooth stellate cells are inhibitory. Pyramidal cells project out of the cortex, whereas both types of stellate cells are local neurons. C. Conception of information flow based on anatomical connections. ( LGN = lateral geniculate nucleus; MT = middle temporal area.) Inputs. Axons from M and P cells in the lateral geniculate nucleus end on spiny stellate cells in the sublayers of 4C, and these cells project axons to layer 4B or the upper layers 2 and 3. Axons from cells in the intralaminar zones of the lateral geniculate nucleus project directly to layers 2 and 3. Intracortical connections. Axon collaterals of pyramidal cells in layers 2 and 3 project to layer 5 pyramidal cells, whose axon collaterals project both to layer 6 pyramidal cells and back to cells in layers 2 and 3. Axon collaterals of layer 6 pyramidal cells then make a loop back to layer 4C onto smooth stellate cells. Output. Each layer, except for 4C, has outputs for V1 and each is different. The cells in layers 2, 3, and 4B project to extrastriate visual cortical areas. Cells in layer 5 project to the superior colliculus, the pons, and the pulvinar. Cells in layer 6 project back to the lateral geniculate nucleus and the claustrum. Layered Structure of the Primary Visual Cortex Like almost all other portions of the cerebral cortex, the primary visual cortex has six distinct layers, as shown in Figure 51–4. Also, as is true for the other sensory systems, the geniculocalcarine fibers terminate mainly in layer IV. But this layer, too, is organized into subdivisions.The rapidly conducted signals from the Y retinal ganglion cells terminate in layer IVca, and from there they are relayed vertically both outward toward the cortical surface and inward toward deeper levels. The visual signals from the medium-sized optic nerve fibers, derived from the X ganglion cells in the retina, also terminate in layer IV, but at points different from the Y signals. They terminate in layers IVa and IVcb, the shallowest and deepest portions of layer IV, shown to the right in Figure 51–4. From there, these signals are transmitted vertically both toward the surface of the cortex and to deeper layers. It is these X ganglion pathways that transmit the accurate pointto- point type of vision as well as color vision. Vertical Neuronal Columns in the Visual Cortex. The visual cortex is organized structurally into several million vertical columns of neuronal cells, each column having a diameter of 30 to 50 micrometers. The same vertical columnar organization is found throughout the cerebral cortex for the other senses as well (and also in the motor and analytical cortical regions). Each column represents a functional unit. One can roughly calculate that each of the visual vertical columns has perhaps 1000 or more neurons. After the optic signals terminate in layer IV, they are further processed as they spread both outward and inward along each vertical column unit. This processing is believed to decipher separate bits of visual information at successive stations along the pathway. The signals that pass outward to layers I, II, and III eventually transmit signals for short distances laterally in the cortex. Conversely, the signals that pass inward to layers V and VI excite neurons that transmit signals much greater distances. “ Color Blobs” in the Visual Cortex. Interspersed among the primary visual columns as well as among the columns of some of the secondary visual areas are special column-like areas called color blobs. They receive lateral signals from adjacent visual columns and are activated specifically by color signals. Therefore, it is presumed that these blobs are the primary areas for deciphering color. Interaction of Visual Signals from the Two Separate Eyes. Recall that the visual signals from the two separate eyes are relayed through separate neuronal layers in the lateral geniculate nucleus. These signals still remain separated from each other when they arrive in layer IV of the primary visual cortex. In fact, layer IV is interlaced with stripes of neuronal columns, each stripe about 0.5 millimeter wide; the signals from one eye enter the columns of every other stripe, alternating with signals from the second eye. This cortical area deciphers whether the respective areas of the two visual images from the two separate eyes are “in register” with each other—that is, whether corresponding points from the two retinas fit with each other. In turn, the deciphered information is used to adjust the directional gaze of the separate eyes so that they will fuse with each other (be brought into “register”). The information observed about degree of register of images from the two eyes also allows a person to distinguish the distance of objects by the mechanism of stereopsis. The primary visual cortex in humans is about 2 mm thick and consists of six layers of cells (layers 1-6) between the pial surface and the underlying white matter. The principal layer for inputs from the lateral geniculate nucleus is layer 4, which is further subdivided into four sublayers (sublaminae): 4A, 4B, 4Cα, and 4Cβ. Tracings of resident cells and axonal inputs in the monkey have shown that the M and P cells of the lateral geniculate nucleus terminate in different layers and even in different sublayers. The axons of M cells terminate principally in sublamina 4Cα; the axons of most P cells terminate principally in sublamina 4Cβ (Figure 27- 10A). Thus, the segregation of the parvocellular and magnocellular pathways continues to be maintained at this level of processing. Axons from a third group of cells, located in the intralaminar region of the lateral geniculate nucleus, terminate in layers 2 and 3, where they innervate patches of cells called blobs , a functional grouping that we shall discuss below. These intralaminar cells probably receive their retinal inputs primarily from ganglion cells other than those providing inputs to the M and P cells. These cells might therefore represent another pathway in parallel to the P and M pathways from the retina to the visual cortex, but little is now known about their function. As we have seen in Chapter 17, the cortex contains two basic classes of cells. Pyramidal cells are large and have long spiny dendrites; they are projection neurons whose axons project to other brain regions as well as interconnecting neurons in local areas. Nonpyramidal cells are small and stellate in shape and have dendrites that are either spiny (spiny stellate cells) or smooth (smooth stellates). They are local interneurons whose axons are confined to the primary visual cortex (Figure 27-10B). The pyramidal and spiny stellate cells are excitatory and many use glutamate or aspartate as their transmitters; the smooth stellate cells are inhibitory and many contain γ-aminobutyric acid (GABA). Once afferents from the lateral geniculate nucleus enter the primary visual cortex, information flows systematically from one cortical layer to another, starting with the spiny stellate cells, which predominate in layer 4. The spiny stellate cells distribute the input from the lateral geniculate nucleus to the cortex and the pyramidal cells feed axon collaterals upward and downward to integrate activity within the layers of V1 (Figure 27-10C).
Figure 27-11 Receptive field of a simple cell in the primary visual cortex. The receptive field of a cell in the visual system is determined by recording activity in the cell while spots and bars of light are projected onto the visual field at an appropriate distance from the fovea. The records shown here are for a single cell. Duration of illumination is indicated by a line above each record of action potentials. (Adapted from Hubel and Wiesel 1959 and Zeki 1993.) 1. The cell's response to a bar of light is strongest if the bar of light is vertically oriented in the center of its receptive field. 2. Spots of light consistently elicit weak responses or no response. A small spot in the excitatory center of the field elicits only a weak excitatory response ( a ). A small spot in the inhibitory area elicits a weak inhibitory response ( b ). Diffuse light produces no response ( c ). 3. By using spots of light, the excitatory or “on” areas (+) and inhibitory or “off” areas (-) can be mapped. The map of the responses reveals an elongated “on” area and a surrounding “off” area, consistent with the optimal response of the cell to a vertical bar of light. Simple and Complex Cells Decompose the Outlines of a Visual Image Into Short Line Segments of Various Orientations How is the complexity of the circuitry in the cerebral cortex reflected in the response properties of cortical cells? Hubel, Wiesel, and their colleagues found that most cells above and below layer 4 respond optimally to stimuli that are substantially more complex than those that excite cells in the retina and lateral geniculate nucleus. Their most unexpected finding was that small spots of light—which are so effective in the retina, lateral geniculate nucleus, and in the input layer of the cortex 4C—are much less effective in all other layers of the visual cortex except possibly the blob regions in the superficial layers. Instead, cells respond best to stimuli that have linear properties, such as a line or bar. These cells belong to two major groups, simple and complex. Figure 12.9. Neurons in the primary visual cortex respond selectively to oriented edges. (A) An anesthetized animal is fitted with contact lenses to focus the eyes on a screen, where images can be projected; an extracellualr electrode records the neuronal responses. (B) Neurons in visual cortex typically respond vigorously to a bar of light oriented at a particular angle and weakly—or not at all—to other orientations. The Functional Organization of the Striate Cortex Much in the same way that Stephen Kuffler explored the response properties of individual retinal ganglion cells (see Chapter 11 ), David Hubel and Torsten Wiesel used microelectrode recordings to examine the properties of neurons in more central visual structures. The responses of neurons in the lateral geniculate nucleus were found to be remarkably similar to those in the retina, with a center-surround receptive field organization and selectivity for luminance increases or decreases. However, the small spots of light that were so effective stimulating neurons in the retina and lateral geniculate nucleus were largely ineffective in visual cortex. Instead, most cortical neurons in cats and monkeys were found to respond vigorously to light-dark bars or edges, and only if the bars were presented at a particular range of orientations within the cell's receptive field ( Figure 12.9 ). The responses of cortical neurons are thus tuned to the orientation of edges, much like cone receptors are tuned to the wavelength of light; the peak in the tuning curve (the orientation to which a cell is most responsive) is referred to as the neuron's preferred orientation. By sampling the responses of a large number of single cells, Hubel and Weisel demonstrated that all edge orientations were roughly equally represented in visual cortex. As a result, a given orientation in a visual scene appears to be “encoded” in the activity of a distinct population of orientation-selective neurons . Hubel and Wiesel also found that there are subtly different subtypes within a class of neurons that preferred the same orientation. For example, the receptive fields of some cortical cells, which they called simple cells , were composed of spatially separate “on” and “off” response zones, as if the “on” and “off” centers of lateral geniculate cells that supplied these neurons were arrayed in separate parallel bands. Other neurons, referred to as complex cells , exhibited mixed “on” and “off” responses throughout their receptive field, as if they received their inputs from a number of simple cells. Further analysis uncovered cortical neurons sensitive to the length of the bar of light that was moved across their receptive field, decreasing their rate of response when the bar exceeded a certain length. Still other cells responded selectively to the direction in which an edge moved across their receptive field. Although the mechanisms responsible for generating these selective responses are still not well understood, there is little doubt that the specificity of the receptive field properties of neurons in the striate cortex (and beyond) plays an important role in determining the basic attributes of visual scenes. Another feature that distinguishes the responses of neurons in the striate cortex from those at earlier stages in the primary visual pathway is binocularity . Although the lateral geniculate nucleus receives inputs from both eyes, the axons terminate in separate layers, so that individual geniculate neurons are monocular, driven by either the left or right eye but not by both ( Figure 12.10 ; see also Figure 12.14 ). In some species, including most (but not all) primates, inputs from the left and right eyes remain segregated to some degree even beyond the geniculate because the axons of geniculate neurons terminate in alternating eye-specific columns within cortical layer IV—the so-called ocular dominance columns (see the next section). Beyond this point, the signals from the two eyes are combined at the cellular level. Thus, most cortical neurons have binocular receptive fields, and these fields are almost identical, having the same size, shape, preferred orientation, and roughly the same position in the visual field of each eye. Bringing together the inputs from the two eyes at the level of the striate cortex provides a basis for stereopsis , the special sensation of depth that arises from viewing nearby objects with two eyes instead of one. Because the two eyes look at the world from slightly different angles, objects that lie in front of or behind the plane of fixation project to noncorresponding points on the two retinas. To convince yourself of this fact, hold your hand at arm's length and fixate on the tip of one finger. Maintain fixation on the finger as you hold a pencil in your other hand about half as far away. At this distance, the image of the pencil falls on noncorresponding points on the two retinas and will therefore be perceived as two separate pencils (a phenomenon called double vision, or diplopia ). If the pencil is now moved toward the finger (the point of fixation), the two images of the pencil fuse and a single pencil is seen in front of the finger. Thus, for a small distance on either side of the plane of fixation, where the disparity between the two views of the world remains modest, a single image is perceived; the disparity between the two eye views of objects nearer or farther than the point of fixation is interpreted as depth ( Figure 12.11 ). Although the neurophysiological basis of stereopsis is not understood, some neurons in the striate cortex and in other visual cortical areas have receptive field properties that make them good candidates for extracting information about binocular disparity. Unlike many binocular cells whose monocular receptive fields sample the same region of visual space, these neurons have monocular fields that are slightly displaced (or perhaps differ in their internal organization) so that the cell is maximally activated by stimuli that fall on noncorresponding parts of the retinas. Some of these neurons (so-called far cells ) discharge to disparities beyond the plane of fixation, while others ( near cells ) respond to disparities in front of the plane of fixation. The pattern of activity in these different classes of neurons seems likely to contribute to sensations of stereoscopic depth ( Box B ). Interestingly, the preservation of the binocular responses of cortical neurons is contingent on the normal activity from the two eyes during early postnatal life. Anything that creates an imbalance in the activity of the two eyes—for example, the clouding of one lens or the abnormal alignment of the eyes during infancy (strabismus)—can permanently reduce the effectiveness of one eye in driving cortical neurons, and thus impair the ability to use binocular information as a cue for depth. Early detection and correction of visual problems is therefore essential for normal visual function in maturity (see Chapter 24 ).
Figure 27-12 The receptive fields of simple cells in the primary visual cortex are different and more varied than those of the neurons in the retina and lateral geniculate nucleus. A. Cells of the retina and lateral geniculate nucleus fall into two classes: on-center and offcenter. The receptive fields of these neurons have a center-surround organization due to antagonistic excitatory (+) and inhibitory (-) regions. B. The receptive fields of simple cells in the primary visual cortex have narrow elongated zones with either excitatory (+) or inhibitory (-) flanking areas. Despite the variety, the receptive fields of simple cells share three features: (1) specific retinal position, (2) discrete excitatory and inhibitory zones, and (3) a specific axis of orientation. C. Model of the organization of inputs in the receptive field of simple cells proposed by Hubel and Wiesel. According to this model, a simple cortical neuron in the primary visual cortex receives convergent excitatory connections from three or more on-center cells that together represent light falling along a straight line in the retina. As a result, the receptive field of the simple cortical cell has an elongated excitatory region, indicated by the colored outline in the receptive field diagram. The inhibitory surround of the simple cortical cells is probably provided by off-center cells whose receptive fields (not shown) are adjacent to those of the on-center cells. (Adapted from Hubel and Wiesel 1962). Figure 27-13 The receptive field of a complex cell in the primary visual cortex has no clearly excitatory or inhibitory zones. Orientation of the light stimulus is important, but position within the receptive field is not. (Adapted from Hubel and Wiesel 1962). A. In this example the cell responds best to a vertical edge moving across the receptive field from left to right. This figure shows the patterns of action potentials fired by the cell in response to two types of variation in the stimulus: differences in orientation and differences in position. The line above each record indicates the period of illumination. Different orientations of the light stimulus produce different rates of firing in the cell. A vertical bar of light on the left of the receptive field produces a strong excitatory response (a). Orientations other than vertical are less effective ( b-d ). The position of the border of the light within the receptive field affects the type of response in the cell. If the edge of the light comes from any point on the right within the receptive field, the stimulus produces an excitatory response ( a-d ). If the edge comes from the left, the stimulus produces an inhibitory response ( f-i ). Illumination of the entire receptive field produces no response ( e ). B. According to Hubel and Wiesel, the receptive fields of complex cells are determined by the pattern of inputs. Each complex cell receives convergent excitatory input from several simple cortical cells, each of which has a receptive field with the same organization: a central rectilinear excitation zone (+) and flanking inhibitory regions (-). In this way the receptive field of a complex cell is built up from the individual fields of the presynaptic cells. The simple cells respond best to a bar of light with a specific orientation. For example, a cell that responds best to a vertical bar will not respond, or respond only weakly, to a bar that is horizontal or even oblique (Figure 27-11). Thus, an array of cells in the cortex, all receiving impulses from the same point on the retina but with rectilinear receptive fields with different axes of orientation, is able to represent every axis of rotation for that point on the retina. Simple cells also have excitatory and inhibitory zones in their receptive fields, although these zones are slightly larger than those for lateral geniculate cells (Figure 27-12A, B). For example, a cell may have a rectilinear excitatory zone (with its long axis running from 12 to 6 o'clock such as in Figure 27-12B upper right). For a cell with such a field, an effective stimulus must excite the specific segment of the retina innervated by receptors in the excitatory zone and have the correct linear properties (in this case an edge) and have a specific axis of orientation (in this case vertical, running from 12 to 6 o'clock). Rectilinear receptive fields could be built up from many circular fields if the presynaptic connections from the lateral geniculate nucleus were appropriately arrayed on the simple cell (Figure 27-12C). Indeed, experiments have indicated that the excitatory (“on”) regions in the receptive field of simple cells largely represent the input from on-center lateral geniculate cells while the inhibitory (“off”) regions represent inputs from off- center lateral geniculate cells. The receptive fields of complex cells in the cortex are usually larger than those of simple cells. These fields also have a critical axis of orientation, but the precise position of the stimulus within the receptive field is less crucial because there are no clearly defined on or off zones (Figure 27-13A). Thus, movement across the receptive field is a particularly effective stimulus for certain complex cells. Although some complex cells have direct connections with cells of layer 4C, Hubel and Wiesel proposed that a significant input to complex cells comes from a group of simple cortical cells with the same axis of orientation but with slightly offset receptive field positions (Figure 27-13B). Some Feature Abstraction Is Accomplished by Progressive Convergence The pattern of convergence of inputs throughout the pathway that leads to the complex cells suggests that each complex cell surveys the activity of a group of simple cells, each simple cell surveys the activity of a group of geniculate cells, and each geniculate cell surveys the activity of a group of retinal ganglion cells. The ganglion cells survey the activity of bipolar cells that, in turn, survey an array of receptors. At each level each cell has a greater capacity for abstraction than cells At each level of the afferent pathway the stimulus properties that activate a cell become more specific. Retinal ganglion and geniculate neurons respond primarily to contrast. This elementary information is transformed in the simple and complex cells of the cortex, through the pattern of excitation in their rectilinear fields, into relatively precise line segments and boundaries. Hubel and Wiesel suggest that this processing is an important step in analyzing the contours of objects. In fact, contour information may be sufficient to recognize an object. Monotonous interior or background surfaces contain no critical visual information! David Hubel describes this unexpected feature of perception: Many people, including myself, still have trouble accepting the idea that the interior of a form… does not itself excite cells in our brain,… that our awareness of the interior as black or white. … depends only on cells' sensitivity to the borders. The intellectual argument is that perception of an evenly lit interior depends on the activation of cells having fields at the borders and on the absence of activation of cells whose fields are within the borders, since such activation would indicate that the interior is not evenly lit. So our perception of the interior as black, white, gray or green has nothing to do with cells whose fields are in the interior—hard as that may be to swallow.… What happens at the borders is the only information you need to know: the interior is boring. It is the information carried by edges that allows us to recognize objects in a picture readily even when the objects are sketched only in rough outline (see Figure 25-3). Since simple and complex cells in V1 receive input from both the M and P pathways, both pathways could contribute to what the theoretical biologist David Marr called the primal sketch , the initial two-dimensional approximation of the shape of a stimulus. We will return in Chapter 28 to the fate of the P and M pathways.
This type of wiring could produce a simple-cell receptive field. On the right, four cells are shown making excitatory synaptic connections with a cell of higher order. Each of the lower-order cells has a radially symmetric receptive field with on- center and off-surround, illustrated by the left side of the diagram. The centers of these fields lie along a line. If we suppose that many more than four center-surround cells are connected with the simple cell, all with their field centers overlapped along this line, the receptive field of the simple cell will consist of a long, narrow excitatory region with inhibitory flanks. Avoiding receptive-field terminology, we can say that stimulating with a small spot anywhere in this long, narrow rectangle will strongly activate one or a few of the center-surround cells and in turn excite the simple cell, although only weakly. Stimulating with a long, narrow slit will activate all the center-surround cells, producing a strong response in the simple cell.
This wiring diagram would account for the properties of a complex cell. As in the figure on page 15, we suppose that a large number of simple cells (only three are shown here) make excitatory synapses with a single complex cell. Each simple cell responds optimally to a vertically oriented edge with light to the right, and the receptive fields are scattered in overlapping fashion throughout the rectangle. An edge falling anywhere within the rectangle evokes a response from a few simple cells, and this in turn evokes a response in the complex cell. Because there is adaptation at the synapses, only a moving stimulus will keep up a steady bombardment of the complex cell.
Neuronal Patterns of Stimulation During Analysis of the Visual Image Analysis of Contrasts in the Visual Image. If a person looks at a blank wall, only a few neurons in the primary visual cortex will be stimulated, regardless of whether the illumination of the wall is bright or weak. Therefore, the question must be asked, What does the primary visual cortex detect? To answer this, let us now place on the wall a large solid cross, as shown to the left in Figure 51–5. To the right is shown the spatial pattern of the most excited neurons in the visual cortex. Note that the areas of maximum excitation occur along the sharp borders of the visual pattern. Thus, the visual signal in the primary visual cortex is concerned mainly with contrasts in the visual scene, rather than with noncontrasting areas. We saw in Chapter 50 that this is true of most of the retinal ganglion cells as well, because equally stimulated adjacent retinal receptors mutually inhibit one another. But at any border in the visual scene where there is a change from dark to light or light to dark, mutual inhibition does not occur, and the intensity of stimulation of most neurons is proportional to the gradient of contrast — that is, the greater the sharpness of contrast and the greater the intensity difference between light and dark areas, the greater the degree of stimulation. Visual Cortex Also Detects Orientation of Lines and Borders— “Simple” Cells. The visual cortex detects not only the existence of lines and borders in the different areas of the retinal image but also the direction of orientation of each line or border—that is, whether it is vertical or horizontal or lies at some degree of inclination. This is believed to result from linear organizations of mutually inhibiting cells that excite second-order neurons when inhibition occurs all along a line of cells where there is a contrast edge. Thus, for each such orientation of a line, specific neuronal cells are stimulated. A line oriented in a different direction excites a different set of cells.These neuronal cells are called simple cells. They are found mainly in layer IV of the primary visual cortex. Detection of Line Orientation When a Line Is Displaced Laterally or Vertically in the Visual Field—“Complex” Cells. As the visual signal progresses farther away from layer IV, some neurons respond to lines that are oriented in the same direction but are not position-specific. That is, even if a line is displaced moderate distances laterally or vertically in the field, the same few neurons will still be stimulated if the line has the same direction. These cells are called complex cells. Detection of Lines of Specific Lengths, Angles, or Other Shapes. Some neurons in the outer layers of the primary visual columns, as well as neurons in some secondary visual areas, are stimulated only by lines or borders of specific lengths, by specific angulated shapes, or by images that have other characteristics. That is, these neurons detect still higher orders of information from the visual scene. Thus, as one goes farther into the analytical pathway of the visual cortex, progressively more characteristics of each visual scene are deciphered. Detection of Color Color is detected in much the same way that lines are detected: by means of color contrast. For instance, a red area is often contrasted against a green area, a blue area against a red area, or a green area against a yellow area. All these colors can also be contrasted against a white area within the visual scene. In fact, this contrasting against white is believed to be mainly responsible for the phenomenon called “color constancy”; that is, when the color of an illuminating light changes, the color of the “white” changes with the light, and appropriate computation in the brain allows red to be interpreted as red even though the illuminating light has changed the color entering the eyes. The mechanism of color contrast analysis depends on the fact that contrasting colors, called “opponent colors,” excite specific neuronal cells. It is presumed that the initial details of color contrast are detected by simple cells, whereas more complex contrasts are detected by complex and hypercomplex cells. Effect of Removing the Primary Visual Cortex Removal of the primary visual cortex in the human being causes loss of conscious vision, that is, blindness. However, psychological studies demonstrate that such “blind” people can still, at times, react subconsciously to changes in light intensity, to movement in the visual scene, or, rarely, even to some gross patterns of vision. These reactions include turning the eyes, turning the head, and avoidance.This vision is believed to be subserved by neuronal pathways that pass from the optic tracts mainly into the superior colliculi and other portions of the older visual system.
The technique of optical imaging. A sensitive video camera is used to record changes in light absorption that occur as the animal views various stimuli presented on a video monitor. Images are digitized and stored in a computer in order to construct maps that compare patterns of activity associated with different stimuli. (B) Maps of orientation preference in the visual cortex visualized with optical imaging. Each color represents the angle of an edge that was most effective in activating the neurons at a given site. Orientation preference changes in a continuous fashion, rotating around pinwheel centers. (C) Comparison of optical image maps of orientation preference and ocular dominance in monkey visual cortex. The thick black lines represent the borders between ocular dominance columns. The thin gray lines represent the iso-orientation contours, which converge at orientation pinwheel centers (arrow). Iso-orientation contour lines generally intersect the borders of ocular dominance bands at right angles. (B from Bonhoeffer and Grinvald, 1993; C from Obermeyer and Blasdel, 1993.) Optical Imaging of Functional Domains in the Visual Cortex The recent availability of optical imaging techniques has made it possible to visualize how response properties, such as the selectivity for edge orientation or ocular dominance, are mapped across the cortical surface. These methods generally rely on intrinsic signals (changes in the amount of light reflected from the cortical surface) that correlate with levels of neural activity. Such signals are thought to arise at least in part from local changes in the ratio of oxyhemoglobin and deoxyhemoglobin that accompany such activity, more active areas having a higher deoxyhemoglobin/oxyhemoglobin ratio (see also Box D in Chapter 1 ). This change can be detected when the cortical surface is illuminated with red light (605–700 nm). Under these conditions, active cortical regions absorb more light than less active ones. With the use of a sensitive video camera, and averaging over a number of trials (the changes are small, 1 or 2 parts per thousand), it is possible to visualize these differences and use them to map cortical patterns of activity (figure A). This approach has now been successfully applied to both striate and extrastriate areas in both experimental animals and human patients undergoing neurosurgery. The results emphasize that maps of stimulus features are a general principle of cortical organization. For example, orientation preference is mapped in a continuous fashion such that adjacent positions on the cortical surface tend to have only slightly shifted orientation preferences. However, there are points where continuity breaks down. Around these points, orientation preference is represented in a radial pattern resembling a pinwheel, covering the whole 180° of possible orientation values (figure B). This powerful technique can also be used to determine how maps for different stimulus properties are arranged relative to one another, and to detect additional maps such as that for direction of motion. A comparison of ocular dominance bands and orientation preference maps, for example, shows that pinwheel centers are generally located in the center of ocular dominance bands, and that the iso-orientation contours that emanate from the pinwheel centers run orthogonal to the borders of ocular dominance bands (figure C). An orderly relationship between maps of orientation selectivity and direction selectivity has also been demonstrated. These systematic relationships between the functional maps that coexist within primary visual cortex are thought to ensure that all combinations of stimulus features (orientation, direction, ocular dominance, and spatial frequency) are analyzed for all regions of visual space. Figure 27-14 Orientation columns in the visual cortex of the monkey. (Courtesy of Gary Blasdel.) Image of a 9 by 12 mm rectangle of cortical surface taken while the monkey viewed contours of different orientations (indicated on the right). This image was obtained through optical imaging and by comparing local changes in reflectance, which indicate activity. Areas that were most active during the presentation of a particular orientation are indicated by the color chosen to represent that orientation (bars on the right). Complementary colors were chosen to represent orthogonal orientations. Hence, red and green indicate maximal activities in response to horizontal and vertical, while blue and yellow indicate greatest activation by left and right oblique. Enlargement of a pinwheel-like area in A. Orientations producing the greatest activity remain constant along radials, extending outward from a center, but change continuously (through ± 18°). C. Three-dimensional organization of orientation columns in a 1 mm × 1 mm × 2 mm slab of primary visual cortex underlying the square surface region depicted in B. The Primary Visual Cortex Is Organized Into Functional Modules We have seen how the organization of the receptive fields of neurons in the visual pathway changes from concentric to simple to complex. Do these local transformations reflect a larger organization within the visual cortex? We shall see that the neurons in the visual cortex have a columnar organization, like the somatic sensory cortex, and that sets of columns can be regarded as functional modules, each of which processes visual information from a specific region of the visual field. Neurons With Similar Receptive Fields Are Organized in Columns Like the somatic sensory cortex, the primary visual cortex is organized into narrow columns of cells, running from the pial surface to the white matter. Each column is about 30 to 100 μm wide and 2 mm deep, and each contains cells in layer 4C with concentric receptive fields. Above and below are simple cells whose receptive fields monitor almost identical retinal positions and have identical axes of orientation. For this reason these groupings are called orientation columns. Each orientation column also contains complex cells. The properties of these complex cells can most easily be explained by postulating that each complex cell receives direct connections from the simple cells in the column. Thus, columns in the visual system seem to be organized to allow local interconnection of cells, from which the cells are able to generate a new level of abstraction of visual information. For instance, the columns allow cortical cells to generate linear receptive field properties from the inputs of several cells in the lateral geniculate nucleus that respond best to small spots of light. The discovery of columns in the various sensory systems was one of the most important advances in cortical physiology in the past several decades and immediately raised questions that have led to a family of new discoveries. For example, given that cells with the same axis of orientation tend to be grouped into columns, how are columns of cells with different axes of orientation organized in relation to one another? Detailed mapping of adjacent columns by Hubel and Wiesel, using tangential penetrations with microelectrodes, revealed a precise organization with an orderly shift in axis of orientation from one column to the next. About every three quarters of a millimeter contained a complete cycle of orientation changes. The anatomical layout of the orientation columns was first demonstrated in electrophysiological experiments in which marks were made in the cortex near the cells that are activated by stimuli at a given orientation. Later, this anatomical arrangement was delineated by injecting 2-deoxyglucose, a glucose analog that can be radio labeled and injected into the brain. Cells that are metabolically active take up the label and can then be detected when sections of cortex are overlaid with x-ray film. Thus, when a stimulus of lines with a given orientation is presented, an orderly array of active and inactive stripes of cells is revealed. A remarkable advance now allows the different orientation columns to be visualized directly in the living cortex. Using either a voltage-sensitive dye or inherent differences in the light scattering of active and inactive cells, a highly sensitive camera can detect the pattern of active and inactive orientation columns during presentation of a bar of light with a specific axis of orientation (Figure 27-14). The systematic shifts in axis of orientation from one column to another is occasionally interrupted by blobs , the peg-shaped regions of cells prominent in layers 2 and 3 of V1 (Figure 27-15). The cells in the blobs frequently respond to different color stimuli, and their receptive fields, like those of cells in the lateral geniculate nucleus, have no specific orientation.
Figure 27-15 Organization of blobs in the visual cortex. A. Blobs are visible as dark patches in this photograph of a single 40μm thick layer of upper cortex that has been processed histochemically to reveal the density of cytochrome oxidase, a mitochondrial enzyme involved in energy production. The heightened enzymatic activity in the blobs is thought to represent heightened neural activity. The cortex was sectioned tangentially. (Courtesy of D. Ts'o, C. Gilbert, and T. Wiesel.) B. Organization of the blobs in relation to the orientation columns. Only the upper layers of the cortex are shown with the blobs extending though these layers. The blobs interrupt the pattern of the orientation columns. Figure 27-16 The ocular dominance columns. A. This autoradiograph of the primary visual cortex of an adult monkey shows the ocular dominance columns as alternating white and dark (labeled and unlabeled) patches in layer 4 of the cortex, below the pial surface. One eye of the monkey was injected with a cell label, which over the course of 2 weeks was transported to the lateral geniculate nucleus and then across synapses to the geniculocortical relay cells, whose axons terminate in layer 4 of the visual cortex. Areas of layer 4 that receive input from the injected eye are heavily labeled and appear white; the alternating unlabeled patches receive input from the uninjected eye. In all, some 56 columns can be counted in layer 4C. The underlying white matter appears white because it contains the labeled axons of geniculate cells. (From Hubel and Wiesel 1979.) B. The scheme of inputs to the alternating ocular dominance columns in layer 4 of the primary visual cortex. Inputs from the contralateral ( C ) and ipsilateral ( I ) eyes arise in different layers in the lateral geniculate nucleus ( LGN ), identified in Figure 27-5, and project to different subdivisions of layer 4. In addition to columns of cells responsive to axis of orientation and blobs related to color processing, a third system of alternating columns processes separate inputs from each eye. These ocular dominance columns, which we shall consider again in Chapter 56, represent an orderly arrangement of cells that receive inputs only from the left or right eye and are important for binocular interaction. The ocular dominance columns have been visualized using transsynaptic transport of radiolabeled amino acids injected into one eye. In autoradiographs of sections of cortex cut perpendicular to the layers, patches in layer 4 that receive input from the injected eye are heavily labeled, and they alternate with unlabeled patches that mediate input from the uninjected eye (Figure 27-16).
Figure 27-17 Organization of orientation columns, ocular dominance columns, and blobs in primary visual cortex. A. An array of functional columns of cells in the visual cortex contains the neural machinery necessary to analyze a discrete region of the visual field and can be thought of as a functional module. Each module contains one complete set of orientation columns, one set of ocular dominance columns (right and left eye), and several blobs (regions of the cortex associated with color processing). The entire visual field can be represented in the visual cortex by a regular array of such modules. B. Images depicting ocular dominance columns, orientation columns, and blobs from the same region of primary visual cortex. (Courtesy of Gary Blasdel.) 1. Images of ocular dominance columns were obtained using optical imaging and independently stimulating the left and right ocular dominance columns in a particular region. Because neural activity decreases cortical reflectance, the subtraction of one left eye image from one right eye image produces the characteristic pattern of dark and light bands, representing the right and left eyes respectively. 2. In this image the borders of the ocular dominance columns shown in 1 appear as black lines superimposed on the pattern of orientation-specific columns depicted in Figure 27-14. 3. The borders of the ocular dominance columns shown in 1 are superimposed on tissue reacted for cytochrome oxidase, which visualizes the blobs. The blobs are thus seen localized in the centers of the ocular dominance columns. A Hypercolumn Represents the Visual Properties of One Region of the Visual Field Hubel and Wiesel introduced the term hypercolumn to refer to a set of columns responsive to lines of all orientations from a particular region in space. The relationship between the orientation columns, the independent ocular dominance columns, and the blobs within a module is illustrated in Figure 27-17. A complete sequence of ocular dominance columns and orientation columns is repeated regularly and precisely over the surface of the primary visual cortex, each occupying a region of about 1 mm2. This repeating organization is a striking illustration of the modular organization characteristic of the cerebral cortex. Each module acts as a window on the visual field and each window represents only a tiny part of the visual field, but the whole field is covered by many such windows. Within the processing module all information about that part of the visual world is processed. From what we know now, that includes orientation, binocular interaction, color, and motion. Each module has a variety of outputs originating in different cortical layers. The organization of the output connections from the primary visual cortex is similar to that of the somatic sensory cortex in that there are outputs from all layers except 4C, and in each layer the principal output cells are the pyramidal cells (see Figure 27-10C). The axons of cells above layer 4C project to other cortical areas; those of cells below 4C project to subcortical areas. The cells in layers 2 and 3 send their output to other higher visual cortical regions, such as Brodmann's area 18 (V2, V3, and V4). They also make connections via the corpus callosum to anatomically symmetrical cortical areas on the other side of the brain. Cells in layer 4B project to the middle temporal area (V5 or MT). Cells in layer 5 project to the superior colliculus, the pons, and the pulvinar. Cells in layer 6 project back to the lateral geniculate nucleus and to the claustrum. Since cells in each layer of the visual cortex probably perform a different task, the laminar position of a cell determines its functional properties.
Figure 27-18 Columns of cells in the visual cortex with similar function are linked through horizontal connections. A. A camera lucida reconstruction of a pyramidal cell injected with horseradish peroxidase in layers 2 and 3 in a monkey. Several axon collaterals branch off the descending axon near the dendritic tree and in three other clusters (arrows). The clustered collaterals project vertically into several layers at regular intervals, consistent with the sequence of functional columns of cells. (From McGuire et al. 1991.) B. The horizontal connections of a pyramidal cell, such as that shown in A, are functionally specific. The axon of the pyramidal cell forms synapses on other pyramidal cells in the immediate vicinity as well as pyramidal cells some distance away. Recordings of cell activity demonstrate that the axon makes connections only with cells that have the same functional specificity (in this case, responsiveness to a vertical line). (Adapted from Ts'o et al. 1986.) C. 1. A section of cortex labeled with 2-deoxyglucose shows a pattern of stripes representing columns of cells that respond to a stimulus with a particular orientation. 2. Microbeads injected into the same site as in 1 are taken up by the terminals of neurons and transported to the cell bodies. 3. Superimposition of the images in 1 and 2. The clusters of bead-labeled cells lie directly over the 2-deoxyglucose-labeled areas, showing that groups of cells in different columns with the same axis of orientation are connected. (From Gilbert and Wiesel 1989.) Columnar Units Are Linked by Horizontal Connections As we have seen, three major vertically oriented systems crossing the layers of primary visual cortex have been delineated: (1) orientation columns, which contain the neurons that respond selectively to light bars with specific axes of orientation; (2) blobs, peg-shaped patches in upper layers (but not layer 4) that contain cells that are more concerned with color than orientation; and (3) ocular dominance columns, which receive inputs from one or the other eye. These units are organized into hypercolumns that monitor small areas of the visual field. These vertically oriented systems communicate with one another by means of horizontal connections that link cells within a layer. Axon collaterals of individual pyramidal cells in layers 3 and 5 run long distances, parallel with the layers, and give rise to clusters of axon terminals at regular intervals that approximate the width of a hypercolumn (Figure 27-18A). Horseradish peroxidase injected into focal regions within superficial cortical layers (2, 3) reveals an elaborate lattice of labeled cells and axons that encloses unlabeled patches about 500 μm in diameter. Similarly, tracers injected into sites corresponding to blobs label other blobs, producing a honeycomb image. A honeycomb array also appears after labeling the nonblob cortex. To examine these horizontal connections, recordings were made from pairs of cells in blob regions; each pair was separated by about 1 mm, the distance that typically separates the lattice arrays described above (Figure 27-18B). Many cell pairs were found to fire simultaneously in response to stimuli with a specific orientation and direction of movement. Thus, colorselective cells in one blob are linked to cells with similar responses in other blobs. Additional evidence that horizontal connections tie together cells with similar response properties in different columns comes from injection of radiolabeled 2-deoxyglucose and fluorescently labeled microbeads into a column containing cells that respond to a specific orientation. The beads are taken up by axon terminals at the injection site and transported back to the cell bodies. In sections tangential to the pia the overall pattern of cells labeled with the microbeads closely resembles the lattice described above. In fact, the pattern labeled with 2-deoxyglucose is congruent with the pattern obtained with the microbeads (Figure 27-18C). Thus, both anatomical and metabolic studies establish that cortical cells having receptive fields with the same orientation are connected by means of a horizontal network. The visual cortex, then, is organized functionally into two sets of intersecting connections, one vertical, consisting of functional columns spanning the different cortical layers, and the other horizontal, connecting functional columns with the same response properties. What is the functional importance of the horizontal connections? Recent studies indicate that these connections integrate information over many millimeters of cortex. As a result, a cell can be influenced by stimuli outside its normal receptive field. Indeed, a cell's axis of orientation is not completely invariant but is dependent on the context on which the feature is embedded. The psycho-physical principle of contextual effect , whereby we evaluate objects in the context in which we see them, is thought to be mediated by the horizontal connections between the functional columns of the visual cortex.
Figure 28-1 Organization of V1 and V2. A. Subregions in V1 (area 17) and V2 (area 18). This section from the occipital lobe of a squirrel monkey at the border of areas 17 and 18 was reacted with cytochrome oxidase. The cytochrome oxidase stains the blobs in V1 and the thick and thin stripes in V2. (Courtesy of M. Livingstone.) B. Connections between V1 and V2. The blobs in V1 connect primarily to the thin stripes in V2, while the interblobs in V1 connect to interstripes in V2. Layer 4B projects to the thick stripes in V2 and to the middle temporal area ( MT ). Both thin and interstripes project to V4. Thick stripes in V2 also project to MT. The Parvocellular and Magnocellular Pathways Feed Into Two Processing Pathways in Extrastriate Cortex In Chapter 27 we saw that the parallel parvocellular and magnocellular pathways remain segregated even in the striate cortex. What happens to these P and M pathways beyond the striate cortex? Early research on these pathways indicated that the P pathway continues in the ventral cortical pathway that extends to the inferior temporal cortex, and that the M pathway becomes the dorsal pathway that extends to the posterior parietal cortex. However, the actual relationships are probably not so exclusive. The evidence for separation of function of the dorsal and ventral pathways begins in the primary visual, or striate, cortex (V1). Staining for the mitochondrial enzyme cytochrome oxidase reveals a precise and repeating pattern of dark, peg-like regions about 0.2 mm in diameter called blobs. The blobs are especially prominent in the superficial layers 2 and 3, where they are separated by intervening regions that stain lighter, the interblob regions. The same stain also reveals alternating thick and thin stripes separated by interstripes of little activity (Figure 28-1 in the secondary visual cortex, or V2).
Figure 28-2 The magnocellular (M) and parvocellular (P) pathways from the retina project through the lateral geniculate nucleus (LGN) to V1. Separate pathways to the temporal and parietal cortices course through the extrastriate cortex beginning in V2. The connections shown in the figure are based on established anatomical connections, but only selected connections are shown and many cortical areas are omitted (compare Figure 25- 9). Note the cross connections between the two pathways in several cortical areas. The parietal pathway receives input from the M pathway but only the temporal pathway receives input from both the M and P pathways. (Abbreviations: AIT = anterior inferior temporal area; CIT = central inferior temporal area; LIP = lateral intraparietal area; Magno = magnocellular layers of the lateral geniculate nucleus; MST = medial superior temporal area; MT = middle temporal area; Parvo = parvocellular layers of the lateral geniculate nucleus; PIT = posterior inferior temporal area; VIP = ventral intraparietal area.) (Based on Merigan and Maunsell 1993.) Margaret Livingstone and David Hubel identified the anatomical connections between labeled regions in V1 and V2 (Figure 28-1B). They found that the P and M pathways remain partially segregated through V2. The M pathway projects from the magnocellular layers of the lateral geniculate nucleus to the striate cortex, first to layer 4Cα and then to layer 4B. Cells in layer 4B project directly to the middle temporal area (MT) and also to the thick stripes in V2, from which cells also project to MT. Thus, a clear anatomical pathway exists from the magnocellular layers in the lateral geniculate nucleus to MT and from there to the posterior parietal cortex (Figure 28-2). Cells in the parvocellular layers of the lateral geniculate nucleus project to layer 4Cβ in the striate cortex, from which cells project to the blobs and interblobs of V1. The blobs send a strong projection to the thin stripes in V2, whereas interblobs send a strong projection to the interstripes in V2. The thin stripe and interstripe areas of V2 may in turn project to discrete subregions of V4, thus maintaining this separation in the P pathway into V4 and possibly on into the inferior temporal cortex. A pathway from the P cells in the lateral geniculate nucleus to the inferior temporal cortex can therefore also be identified (Figure 28-2). But are these pathways exclusive of each other? Several anatomical observations suggest that they are not. In V1 both the magnocellular and parvocellular pathways have inputs in the blobs, and local neurons make extensive connections between the blob and interblob compartments. In V2 cross connections exist between the stripe compartments. Thus, the separation is not absolute, but whether there is an intermixing of the M and P contributions or whether the cross connections allow one cortical pathway to modulate activity in the other is not clear. Results of experiments that selectively inactivate the P and M pathways as they pass through the lateral geniculate nucleus (described in Chapter 27) also erode the notion of strict segregation between the pathways in V1. Blocking of either pathway affects the responses of fewer than half the neurons in V1, which indicates that most V1 neurons receive physiologically effective inputs from both pathways. Further work has shown that the responses of neurons both within and outside of the blobs in the superficial layers of V1 are altered by blocking only the M pathway. Both observations suggest that there is incomplete segregation of the M and P pathways in V1. This selective blocking of the P and M pathways also reveals the relative contributions of the pathways to the parietal and inferior temporal cortices. Blocking the magnocellular layers of the lateral geniculate nucleus eliminates the responses of many cells in MT and always reduces the responses of the remaining cells; blocking the parvocellular layers produces a much weaker effect on cells in MT. In contrast, blocking the activity of either the parvocellular or magnocellular layers in the lateral geniculate nucleus reduces the activity of neurons in V4. Thus, the dorsal pathway to MT seems primarily to include input from the M pathway, whereas the ventral pathway to the inferior temporal cortex appears to include input from both the M and P pathways. We can now see that there is substantial segregation of the P and M pathways up to V1, probably separation into V2, a likely predominance of the M input to the dorsal pathway to MT and the parietal cortex, and a mixture of P and M input into the pathway leading to the inferior temporal lobe (as indicated by the lines crossing between the pathways in Figure 28-2). What should we conclude about the organization of visual processing throughout the multiple areas of the visual cortex? First, we know that there are specific serial pathways through the multiple visual areas, not just a random assortment of equally connected areas. There is substantial evidence for two major processing pathways, a dorsal one to the posterior parietal cortex and a ventral one to the inferior temporal cortex, but other pathways may also exist. Second, there is strong evidence that the processing in these two cortical pathways is hierarchical. Each level has strong projections to the next level (and projections back), and the type of visual processing changes systematically from one level to the next. Third, the functions of cortical areas in the two cortical pathways are strikingly different, as judged both by the anatomical connections and the cellular activity considered in this chapter and by the behavioral and brain imaging evidence discussed in Chapter 25. Our examination of the functional organization within these vast regions of extrastriate visual cortex begins with the dorsal cortical pathway and the most intensively studied visual attribute, motion. We then examine the processing of depth information in the dorsal pathway. Finally, we turn to the ventral cortical pathway and consider the processing of information related to form. Color vision is the subject of the next chapter.
Figure 28-3 Motion in the visual field can be perceived in two ways. A. When the eyes are held still, the image of a moving object traverses the retina. Information about movement depends upon sequential firing of receptors in the retina. B. When the eyes follow an object, the image of the moving object falls on one place on the retina and the information is conveyed by movement of the eyes or the head. Motion Is Analyzed Primarily in the Dorsal Pathway to the Parietal Cortex We usually think of motion as an object moving in the visual field, a car or a tennis ball, and we easily distinguish these moving objects from the stationary background. However, we often see objects in motion not because they move on our retina, but because we track them with eye movements; the image remains stationary on the retina but we perceive movement because our eyes move (Figure 28-3). Motion in the visual field is detected by comparing the position of images recorded at different times. Since most cells in the visual system are exquisitely sensitive to retinal position and can resolve events separated in time by 10 to 20 milliseconds, most cells in the visual system should, in principle, be able to extract information about motion from the position of the image on the retina by comparing the previous location of an object with its current location. What then is the evidence for a special neural subsystem specialized for motion?
Figure 28-4 The illusion of apparent motion is evidence that the visual system analyzes motion in a separate pathway. A. Actual motion is experienced as a sequence of visual sensations, each resulting from the image falling on a different position in the retina. B. Apparent motion may actually be more convincing than actual motion, and is the perceptual basis for motion pictures. Thus, when two lights at positions 1 and 2 are turned on and off at suitable intervals, we perceive a single light moving between the two points. This perceptual illusion cannot be explained by processing of information based on different retinal positions and is therefore evidence for the existence of a special visual system for the detection of motion. (From Hochberg 1978.) The initial evidence for a special mechanism designed to detect motion independent of retinal position came from psychophysical observations on apparent motion , an illusion of motion that occurs when lights separated in space are turned on and off at appropriate intervals (Figure 28-4). The perception of motion of objects that in fact have not changed position suggests that position and motion are signaled by separate pathways.
Figure 28-5 Separate human brain areas are activated by motion and color. Motion studies. Six subjects viewed a black and white random-dot pattern that moved in one of eight directions or remained stationary. The figure shows the effect of motion because the PET scans taken while the pattern was stationary were subtracted from those taken while the pattern was moving. The white and red areas show the high end of activity (increased blood flow). The areas are located on the convexity of the prestriate cortex at the junction of Brodmann's areas 19 and 37. Color studies. The subjects viewed a collage of 15 squares and rectangles of different colors or, alternatively, the same patterns in gray shades only. The figure shows the difference in blood flow while viewing the color and gray patterns. The area showing increased flow, subserving the perception of color, is located inferiorly and medially in the occipital cortex. (From Zeki et al. 1991). Motion Is Represented in the Middle Temporal Area Experiments on monkeys show that neurons in the retina and lateral geniculate nucleus, as well as many areas in the striate and extrastriate cortex, respond very well to a spot of light moving across their receptive fields. In area V1, however, cells respond to motion in one direction, while motion in the opposite direction has little or no effect on them. This directional selectivity is prominent among cells in layer 4B of the striate cortex. Thus, cells in the M pathway provide input to cells in 4B, but these input cells themselves do not show directional selectivity. They simply provide the raw input for the directionally selective cortical cells. In monkeys one area at the edge of the parietal cortex, the middle temporal area (MT), appears to be devoted to motion processing because almost all of the cells are directionally selective and the activity of only a small fraction of these cells is substantially altered by the shape or the color of the moving stimulus. Like V1, MT has a retinotopic map of the contralateral visual field, but the receptive fields of cells within this map are about 10 times wider than those of cells in the striate cortex. Cells with similar directional specificity are organized into vertical columns running from the surface of the cortex to the white matter. Each part of the visual field is represented by a set of columns in which cells respond to different directions of motion in that part of the visual field. This columnar organization is similar to that seen in V1. Cells in MT respond to motion of spots or bars of light by detecting contrasts in luminance. Some cells in MT also respond to moving forms that are not defined by differences in luminance but by differences only in texture or color. While these cells are not selective for color itself, they nonetheless detect motion by responding to an edge defined by color. Thus, even though MT and the dorsal pathway to the parietal cortex may be devoted to the analysis of motion, the cells are sensitive to stimuli (color) that were thought to be analyzed primarily by cells in the ventral pathway. Stimulus information on motion, form, and color therefore is not processed exclusively in separate functional pathways. This description of motion processing is based on research on the MT area in monkeys. In the human brain an area devoted to motion has been identified at the junction of the parietal, temporal, and occipital cortices. Figure 28-5 shows changes in blood flow in this area in PET scans made while the subject viewed a pattern of dots in motion. A cortical area adjacent to MT, the medial superior temporal area (MST), also has neurons that are responsive to visual motion and these neurons may process a type of global motion in the visual field called optic flow, which is important for a person's own movements through an environment (Box 28-1). Box 28-1 Optic Flow Optic flow refers to the perceived motion of the visual field that results from an individual's own movement through the environment. With optic flow the entire visual field moves, in contrast to the local motion of objects. Optic flow provides two types of cues: information about the organization of the environment (near objects will move faster than more distant objects) and information about the control of posture (side-to-side patterns induce body sway). Particularly influential in the development of ideas about optic flow was the demonstration by the experimental psychologist James J. Gibson that optic flow is critical for indicating the direction of observer movement (“heading”). For example, when an individual moves forward with eyes and head directed straight ahead, optic flow expands outward from a point straight ahead in the visual field, a pattern that is frequently used in movies to show space ship flight. Where is optic flow represented in the brain? Neurons in one region of the medial superior temporal area of the parietal cortex in monkeys respond in ways that would make these cells ideal candidates to analyze optic flow. These neurons respond selectively to motion, have receptive fields that cover large parts of the visual field, and respond preferentially to large field motion in the visual field. Additionally, the neurons are sensitive to shifts in the origin of full-field motion and to differences in speed between the center and periphery of the field. The neurons also receive input related to eye movement, which is particularly significant because forward movement is typically accompanied by eye and head movement. Finally, electrical stimulation of this area alters the ability of the monkey to locate the point of origin of field motion, providing further evidence that the superior temporal area of the parietal cortex is important for optic flow.
Figure 28-9 A monkey with an MT lesion and a human patient with damage to extrastriate visual cortex have similar deficits in motion perception. A. Displays used to study the perception of motion. In the display on the left there is no correlation between the directions of movement of several dots, and thus no net motion in the display. In the display on the right all the dots move in the same direction (100% correlation). An intermediate case is in the center; 50% of the dots move in the same direction while the other 50% move in random directions (essentially noise added to the signal). (From Newsome and Pare 1988.) B. The performance of a monkey before and after an MT lesion ( left ). The performance of a human subject with bilateral brain damage is compared to two normal subjects ( right ). The ordinate of the graph shows the percent correlation in the directions of all moving dots (as in part A) required for the monkey to pick out the one common direction. The abscissa indicates the size of the displacement of the dot and thus the degree of apparent motion. Note the general similarity between the performance of the humans and that of the monkey and the devastation to this performance after the cortical lesions. (From Newsome and Pare 1988, Control of Movement Is Selectively Impaired by Lesions of MT These correlations of neuronal activity and visual perception raise the question, Is the activity of direction-selective cells in MT causally related to the visual perception of motion and the control of motion-dependent movement? The question whether direction-selective cells in MT directly affect the control of movement was first addressed in an experiment that examined the relationship of these cells to smooth-pursuit eye movements, the movements that keep a moving target in the fovea (see Figure 28-3). When discrete focal chemical lesions were made within different regions of the retinotopic map in MT of a monkey, the speed of the moving target could no longer be estimated correctly in the region of the visual field monitored by the damaged MT area. In contrast, the lesions did not affect pursuit of targets in other regions of the visual field nor did they affect eye movements to stationary targets. Thus, visual processing in MT is selective for motion of the visual stimulus; lesions produce a blind spot, or a scotoma, for motion. Human patients with lesions of parietal cortex also sometimes have these deficits in smooth pursuit eye movements, but the most frequent behavioral deficit is quite different from that seen after lesions of MT. The neurologist Gordon Holmes originally reported that these patients were unable to follow a target when it was moving toward the side of the brain that had the lesion. For example, a patient with a lesioned right hemisphere has difficulty pursuing a target moving toward the right (Figure 28-8B). Later experiments on monkeys showed that lesions centered on the medial superior temporal area (MST), the next level of processing for visual motion, produced just such a deficit (Figure 28-8A). Perception of Motion Is Altered by Lesions and Microstimulation of MT The question whether MT cells contribute to the perception of visual motion was addressed in an experiment in which monkeys were trained to report the direction of motion in a display of moving dots. The experimenter varied the proportion of dots that moved in the same direction random and at 100% correlation the motion of all dots was in one direction (Figure 28-9A). While normal monkeys could perform the task with less than 10% of the dots moving in the same direction, monkeys with a lesion in MT required nearly 100% coherence to perform as well (Figure 28-9B). A human patient with bilateral brain damage also lost the perception of motion when tested on the same task (Figure 28-9B). In both the monkeys and the human subject, visual acuity for stationary stimuli was not affected by the brain damage. Thus damage to MT reduces the ability of monkeys to detect motion in the visual field, as indicated by disruptions in the pursuit of moving objects and perception of the direction of motion. However, monkeys with MT lesions quickly recover these functions. Directionally selective cells in other areas of cerebral cortex, such as MST, apparently can take over the function performed by MT. Recovery of function is greatly slowed when the lesion affects not only MT but also MST and other extrastriate areas.
Figure 28-11 Monocular depth cues provide information on the relative distance of objects and have been used by painters since antiquity. A. The upper drawing shows the side view of a scene. When the scene is traced on a plane of glass held between the eye and the scene (lower drawing) the resulting two-dimensional tracing reveals the cues needed to perceive depth. Occlusion: The fact that rectangle 4 interrupts the outline of 5 indicates which object is in front, but not how much distance there is between them. Linear perspective: Although lines 6-7 and 8-9 are parallel in reality, they converge in the picture plane. Size perspective: Because the two boys are similiar figures, the smaller boy (2) is assumed to be more distant than the larger boy (1) in the picture plane. Familiar size: The man (3) and the nearest boy are drawn to nearly the same size in the picture. If we know that the man is taller than the boy, we deduce on the basis of their sizes in the picture that the man is more distant than the boy. This type of cue is weaker than the others. (Adapted from Hochberg 1968.) B. Motion of the observer or sideways movement of head and eyes produces depth cues. If the observer moves to the left while looking at the tree, objects closer than the tree move to the right; those farther away move to the left. The full-field motion that results from the observer's own movement is referred to as optic flow. (see Box 28-1.) (Adapted from Busettini et al. 1996). Depth Vision Depends on Monocular Cues and Binocular Disparity One of the major tasks of the visual system is to convert a two-dimensional retinal image into three dimensions. How is this transformation achieved? How do we tell how far one thing is from another? How do we estimate the relative depth of a three-dimensional object in the visual field? Psychophysical studies indicate that the shift from two to three dimensions relies on two types of clues: monocular cues for depth and stereoscopic cues for binocular disparity. Monocular Cues Create Far-Field Depth Perception At distances greater than about 100 feet the retinal images seen by each eye are almost identical, so that looking at a distance we are essentially one-eyed. Nevertheless we can perceive depth with one eye by relying on a variety of tricks called monocular depth cues. Several of these monocular cues were appreciated by the artists of antiquity, rediscovered during the Renaissance, and codified early in the sixteenth century by Leonardo da Vinci. ● Familiar size. If we know from experience something about the size of a person, we can judge the person's distance (Figure 28-11A). ● Occlusion. If one person is partly hiding another person, we assume the person in front is closer (Figure 28-11A). ● Linear perspective Parallel lines, such as those of a railroad track, appear to converge with distance. The greater the convergence of lines, the greater is the impression of distance. The visual system interprets the convergence as depth by assuming that parallel lines remain parallel (Figure 28-11A). ● Size perspective If two similar objects appear different in size, the smaller is assumed to be more distant (Figure 28-11A). ● Distribution of shadows and illumination Patterns of light and dark can give the impression of depth. For example, brighter shades of colors tend to be seen as nearer. In painting this distribution of light and shadow is called chiaroscuro. ● Motion (or monocular movement) parallax. Perhaps the most important of the monocular cues, this is not a static pictorial cue and therefore does not come to us from the study of painting. As we move our heads or bodies from side to side, the images projected by an object in the visual field move across the retina. Objects closer than the object we are looking at seem to move quickly and in the direction opposite to our own movement, whereas more distant objects move more slowly and in the same direction as our movement (Figure 28-11B).
Figure 28-13 Neuronal basis of stereoscopic vision. (Adapted from Ohzawa et al. 1996.) A. When an observer looks at point P the image P′ falls on corresponding points on the retina of each eye. These images completely overlap and therefore have zero binocular disparity. When looking at a point to the left and closer, point Q, the image Q′ in the left eye falls on the same point as P′, but the image in the right eye is laterally displaced. These images have binocular disparity. B. A cortical neuron receiving binocular inputs is maximally activated when the inputs from the two eyes have zero disparity as at P′. C. Another cortical neuron receiving binocular inputs responds best when the inputs from the two eyes are spatially disparate on the two retinas (Q′); it is most sensitive to near stimuli. Information From the Two Eyes Is First Combined in the Primary Visual Cortex How is stereopsis accomplished? Clearly the brain must somehow calculate the disparity between the images seen by the two eyes and then estimate distance based on simple geometric relations. However, this cannot occur before information from the two eyes comes together, and cells in the primary visual cortex (V1) are the first in the visual system to receive input from the two eyes (Chapter 27). Stereopsis, however, requires that the inputs from the two eyes be slightly different —there must be a horizontal disparity in the two retinal images (Figure 28- 13). The important finding that certain neurons in V1 are actually selective for horizontal disparity was made in 1968 by Horace Barlow, Colin Blakemore, Peter Bishop, and Jack Pettigrew. They foundth at a neuron that prefers an oriented bar of light at one place in the visual field responds better when that stimulus appears in front of the screen (referred to as a near stimulus) or when the stimulus is beyond the screen (a far stimulus). There is thus an additional level of organization of information in the ocular dominance columns in V1. Cells sensitive to binocular disparity are found in several cortical visual areas. In addition to V1, some cells in the extrastriate areas V2 and V3 respond to disparity, and many directionselective cells in MT respond best to stimuli at specific distances, either at the plane of fixation or nearer or farther than the plane. Some cells in MST, the next step in the parietal pathway, fire in response to combinations of disparity and direction of motion. That is, the direction of motion preferred by the cell varies with the disparity of the stimulus. For example, a cell that responds to leftward-moving far stimuli might also respond to rightward-moving near stimuli. These cells can convey information not only about the direction of motion but about the direction of motion at different depths within the visual field (as in Figure 28-11B). Studies of cells in the striate and extrastriate cortex that respond selectively to binocular disparity fall into several broad categories. Among these, tuned cells respond best to stimuli at a specific disparity, frequently on the plane of fixation. Other cells respond best to stimuli at a range of disparities either in front of the fixation plane (“near cells”) or beyond the plane (“far cells”) (Figure 28-14) Just as the motion information processed in MT is used both for the visual guidance of movement and for visual perception, disparity-sensitive cells in different regions of visual cortex may use disparity information for different purposes. One use is the perception of depth, which we have already considered. Another is in aligning the eyes to focus at a particular depth in the field. The eyes rotate toward each other (convergence) to focus on near objects and rotate apart (divergence) to focus on more distant objects. The ability to align the eyes develops in the first few months of life and disparity information may play a key role in establishing this alignment. “ Fusion” of the Visual Images from the Two Eyes To make the visual perceptions more meaningful, the visual images in the two eyes normally fuse with each other on “corresponding points” of the two retinas. The visual cortex plays an important role in fusion. It was pointed out earlier in the chapter that corresponding points of the two retinas transmit visual signals to different neuronal layers of the lateral geniculate body, and these signals in turn are relayed to parallel neurons in the visual cortex. Interactions occur between these cortical neurons to cause interference excitation in specific neurons when the two visual images are not “in register”—that is, are not precisely “fused.” This excitation presumably provides the signal that is transmitted to the oculomotor apparatus to cause convergence or divergence or rotation of the eyes so that fusion can be re-established. Once the corresponding points of the two retinas are in register, excitation of the specific “interference” neurons in the visual cortex disappears. Neural Mechanism of Stereopsis for Judging Distances of Visual Objects In Chapter 49, it is pointed out that because the two eyes are more than 2 inches apart, the images on the two retinas are not exactly the same. That is, the right eye sees a little more of the right-hand side of the object, and the left eye a little more of the left-hand side, and the closer the object, the greater the disparity. Therefore, even when the two eyes are fused with each other, it is still impossible for all corresponding points in the two visual images to be exactly in register at the same time. Furthermore, the nearer the object is to the eyes, the less the degree of register. This degree of nonregister provides the neural mechanism for stereopsis, an important mechanism for judging the distances of visual objects up to about 200 feet (60 meters). The neuronal cellular mechanism for stereopsis is based on the fact that some of the fiber pathways from the retinas to the visual cortex stray 1 to 2 degrees on each side of the central pathway.Therefore, some optic pathways from the two eyes are exactly in register for objects 2 meters away; still another set of pathways is in register for objects 25 meters away. Thus, the distance is determined by which set or sets of pathways are excited by nonregister or register. This phenomenon is called depth perception, which is another name for stereopsis.
Figure 28-16 Illusions of edges used to study the higher level information processing in V2 cells of the monkey. A. Examples of illusory contours. 1. A white triangle is clearly seen, although it is not defined in the picture by a continuous border. 2. A vertical bar is seen, although again there is no continuous border. 3. Slight alterations obliterate the perception of the bar seen in 2. 4. The curved contour is not represented by any edges or lines. (From Von der Heydt et al. 1984.) B. A neuron in V2 responds to illusory contours. The cell's receptive field is represented by an ellipse in the drawings on the left. 1. A cell responds to a bar of light moving across its receptive field. Each dot in the record on the right indicates a cell discharge and successive lines indicate the cell's response to successive movements of the bar. 2. The neuron also responds when an illusory contour passes over its receptive field. 3, 4. When only half of the stimulus moves across the cell's receptive field, the response resembles spontaneous activity ( 5 ). (Adapted from Von der Heydt et al. 1984.) Cells in V4 Respond to Form Initial observations on cells in V4 indicated that the cells were selective for color, and it was thought that they were devoted exclusively to color vision. However, many of these same cells are also sensitive to the orientation of bars of light and are more responsive to finer-grained than to coarse-grained stimuli. Thus, some V4 cells are responsive to combinations of color and form. Does removal of V4 alter a monkey's responses to color more than to form? Experiments show that ablation of V4 impairs a monkey's ability to discriminate patterns and shapes but only minimally affects its ability to distinguish colors with different hues and saturation. In other experiments ablation of V4 altered only subtle color discriminations, such as the ability to identify colors under different illumination conditions ( color constancy ). We have noted that some humans lose color vision (achromatopsia) after localized damage to the ventral occipital cortex. PET scans of normal human subjects reveal an increase in activity in the lingual and fusiform gyri when colored stimuli are presented (see Figure 28-5). The deficits in patients with achromatopsia differ from those in monkeys with lesions of V4. The human patients cannot discriminate hues but can discriminate shape and texture, whereas the monkeys' ability to differentiate shapes is markedly diminished while hue discrimination is only minimally affected. It therefore seems unlikely that the area identified in the human brain is directly comparable to the V4 region in the monkey, but instead includes more extended regions, including the inferior temporal cortex, the area we consider next.
Figure 28-17 Many inferior temporal neurons respond both to form and color. A. Average responses for a single neuron to stimuli with different shapes. The height of each bar indicates the average discharge rate during presentation of the stimulus. The dashed line indicates the background discharge rate. B. Responses of the same neuron to colored stimuli. Discharge rates are indicated by the size of each circle. The open circle represents a discharge rate of 30 spikes/s. The responses are plotted on a color map with the relative location of colors, red, green, and blue given for reference. The axes are relative amounts of primary colors. (Adapted from Komatsu and Ideura 1993.)
Figure 28-18 Response of a neuron in the inferior temporal cortex to complex stimuli. The cell responds strongly to the face of a toy monkey ( A ). The critical features producing the response are revealed in a configuration of two black spots and one horizontal black bar arranged on a gray disk ( B ). The bar, spots, and circular outline together were essential, as can be seen by the cell's responses to images missing one or more of these features ( C, D, E, F ). The contrast between the inside and outside of the circular contour was not critical ( G ). However, the spots and bar had to be darker than the background within the outline ( H ). (i = spikes.) (Modified from Kobatake and Tanaka 1994.) Recognition of Faces and Other Complex Forms Depends Upon the Inferior Temporal Cortex We are capable of recognizing and remembering an almost infinite variety of shapes independent of their size or position on the retina. Clinical work in humans and experimental studies in monkeys suggest that form recognition is closely related to processes that occur in the inferior temporal cortex. The response properties of cells in the inferior temporal cortex are those we might expect from an area involved in a later stage of pattern recognition. For example, the receptive field of virtually every cell includes the foveal region, where fine discriminations are made. Unlike cells in the striate cortex and many other extra-striate visual areas, the cells in the inferior temporal area do not have a clear retinotopic organization, and the receptive fields are very large and occasionally may include the entire visual field (both visual hemifields). Such large fields may be related to position invariance , the ability to recognize the same feature anywhere in the visual field. For example, even a small eye movement can easily move an edge stimulus from the receptive field of one V1 neuron to another. In contrast, such a movement would simply move the edge within the receptive field of one inferior temporal neuron. The larger receptive field of many extrastriate regions, including the inferior temporal, may be important in the ability to recognize the same object regardless of its location. The most prominent visual input to the inferior temporal cortex is from V4, so it would not be surprising to see a continuation of the visual processing observed in V4. Inferior temporal cortex appears to have functional subregions and, like V4, may have separate pathways to these regions. Also like V4, inferior temporal cells are sensitive to both shape and color. Many cells in inferior temporal cortex respond to a variety of shapes and colors, although the strength of the response varies for different combinations of shape and color (Figure 28-17). Other cells are selective only for shape or color. Most interesting is the finding that some inferotemporal cells respond only to specific types of complex stimuli, such as the hand or face. For cells that respond to a hand, the individual fingers are a particularly critical visual feature; these cells do not respond when there are no spaces separating the fingers. However, all orientations of the hand elicit similar responses. Among neurons selective for faces, the frontal view of the face is the most effective stimulus for some, while for others it is the side view. Moreover, whereas some neurons respond preferentially to faces, others respond prefer-entially to specific facial expressions. Although the proportion of cells in the inferior temporal cortex responsive to hands or faces is small, their existence, together with the fact that lesions of this region lead to specific deficits in face recognition (Chapter 25), indicates that the inferior temporal cortex is responsible for face recognition. One of the major issues in understanding the brain's analysis of complex objects is the degree to which individual cells respond to the simpler components of these objects. Certain critical elements of faces are sufficient to activate some inferior temporal neurons. For example, instead of a face, two dots and a line appropriately positioned might activate the cell (Figure 28-18). Other experiments suggest that some cells respond to facial dimensions (distance between the eyes) and others to the familiarity of the face. There is also evidence that cells responding to similar features are organized in columns.
Figure 29-1 Color vision enriches visual perception but alone it is a poor detector of spatial detail. (Images courtesy of K. R. Gegenfurtner.) A A normal full-color image contains information about variations in brightness and color. B An achromatic image captures brightness variations in the scene and is formed by weighting the energy of the reflected light by the overall spectral sensitivity of the eye. Spatial detail is easily discerned in this kind of image. C A purely chromatic image contains no information about variations in brightness in the scene; rather it contains only information about hue and saturation. Spatial detail is hard to discern.
Figure 29-3 Context influences the appearance of color. The crosses in the two parts of the figure are printed in the same ink (confirm this by looking at where they are joined at the left, yet they look different because they are surrounded by different backgrounds. The perceived color in the cross tends toward the complement of the background color. (From Albers 1975.)
Figure 11.12. Color vision. The absorption spectra of the four photopigments in the normal human retina. The solid curves indicate the three kinds of cone opsins; the dashed curve shows rod rhodopsin for comparison. Absorbance is defined as the log value of the intensity of incident light divided by intensity of transmitted light. Cones and Color Vision A special property of the cone system is color vision. Perceiving color allows humans (and many other animals) to discriminate objects on the basis of the distribution of the wavelengths of light that they reflect to the eye. While differences in luminance are often sufficient to distinguish objects, color adds another perceptual dimension that is especially useful when differences in luminance are subtle or nonexistent. Color obviously gives us a quite different way of perceiving and describing the world we live in. Unlike rods, which contain a single photo pigment, there are three types of cones that differ in the photopigment they contain. Each of these photo pigments has a different sensitivity to light of different wavelengths, and for this reason are referred to as “blue,” “green,” and “red,” or, more appropriately, short (S), medium (M), and long (L) wavelength cones, terms that more or less describe their spectral sensitivities ( Figure 11.12 ). This nomenclature implies that individual cones provide color information for the wavelength of light that excites them best. In fact, individual cones, like rods, are entirely color blind in that their response is simply a reflection of the number of photons they capture, regardless of the wavelength of the photon (or, more properly, its vibrational energy). It is impossible, therefore, to determine whether the change in the membrane potential of a particular cone has arisen from exposure to many photons at wavelengths to which the receptor is relatively insensitive, or fewer photons at wavelengths to which it is most sensitive. This ambiguity can only be resolved by comparing the activity in different classes of cones. Based on the responses of individual ganglion cells, and cells at higher levels in the visual pathway (see Chapter 12 ), comparisons of this type are clearly involved in how the visual system extracts color information from spectral stimuli. Despite these insights, understanding of the neural mechanisms that underlie color perception has been elusive ( Box D ). Much additional information about color vision has come from studies of individuals with abnormal color detecting abilities. Color vision deficiencies result either from the inherited failure to make one or more of the cone pigments or from an alteration in the absorption spectra of cone pigments (or, rarely, from lesions in the central stations that process color information; see Chapter 12 ). Under normal conditions, most people can match any color in a test stimulus by adjusting the intensity of three superimposed light sources generating long, medium, and short wavelengths. The fact that only three such sources are needed to match (nearly) all the perceived colors is strong confirmation of the fact that color sensation is based on the relative levels of activity in three sets of cones with different absorption spectra. That color vision is trichromatic was first recognized by Thomas Young at the beginning of the nineteenth century (thus, people with normal color vision are called trichromats ). For about 5–6% of the male population in the United States and a much smaller percentage of the female population, however, color vision is more limited. Only two colors of light are needed to match all the colors that these individuals can perceive; the third color category is simply not seen. Such dichromacy , or “color blindness” as it is commonly called, is inherited as a recessive, sex-linked characteristic and exists in two forms: protanopia , in which all color matches can be achieved by using only green and blue light, and deuteranopia , in which all matches can be achieved by using only blue and red light. In another major class of color deficiencies, all three light sources (i.e., short, medium, and long wavelengths) are needed to make all possible color matches, but the matches are made using values that are significantly different from those used by most individuals. Some of these anomalous trichromats require more red than normal to match other colors (protanomalous trichromats); others require more green than normal (deuteranomalous trichromats). Jeremy Nathans and his colleagues at Johns Hopkins University have provided a deeper understanding of these color vision deficiencies by identifying and sequencing the genes that encode the three human cone pigments ( Figure 11.13 ). The genes that encode the red and green pigments show a high degree of sequence homology and lie adjacent to each other on the X chromosome, thus explaining the prevalence of color blindness in males. In contrast, the blue-sensitive pigment gene is found on chromosome 7 and is quite different in its amino acid sequence. These facts suggest that the red and green pigment genes evolved relatively recently, perhaps as a result of the duplication of a single ancestral gene; they also explain why most color vision abnormalities involve the red and green cone pigments. Human dichromats lack one of the three cone pigments, either because the corresponding gene is missing or because it exists as a hybrid of the red and green pigment genes (see Figure 11.13 ). For example, some dichromats lack the green pigment gene altogether, while others have a hybrid gene that is thought to produce a red-like pigment in the “green” cones. Anomalous trichromats also possess hybrid genes, but these elaborate pigments whose spectral properties lie between those of the normal red and green pigments. Thus, although most anomalous trichromats have two distinct sets of long-wavelength cones (one normal, one hybrid), there is more overlap in their absorption spectra than in normal trichromats, and thus less difference in how the two sets of cones respond to a given wavelength (with resulting anomalies in color perception).
Figure 29-4 An object's surface reflectance function describes the fraction of incident light that its surface reflects at each wavelength. A reflectance of 1 means that all the incident light is reflected. The pigments in natural surfaces generally have reflectance functions that vary slowly and smoothly across the visible spectrum, while synthetic pigments often have a more complex structure. A Surface reflectance functions of flowers. ( 1 ) Pale yellow gladiolus reflects little light at short wavelengths but quite uniformly reflects light at wavelengths greater than about 520 nm. ( 2 ) Bright orange gladiolus reflects light mostly at wavelengths greater than 580 nm. ( 3 ) Pale violet rose of Sharon reflects light at long wavelengths and short wavelengths, but not at middle wavelengths (the yellowish-green part of the spectrum). ( 4 ) Wine-colored gladiolus reflects light at long wavelengths and a little at short wavelengths. (From Evans 1948.) B Surface reflectance functions of human skin: white skin; black skin. (From Evans 1948, after Edwards and Duntley 1939.) C Surface reflectance function of light green enamel paint. The surface reflectance functions of synthetic pigments often vary less smoothly with wavelength than do those of natural surfaces. (From Wyszecki and Stiles 1982.)
Figure 29-5 The light reflected from a surface depends both upon the object's surface reflectance function and the spectral composition of the illuminant. The plot shows the spectral composition of light reflected from the surface of a blue vase illuminated by direct sunlight ( 1 ) and by a uniformly overcast sky ( 2 ). The different phases of daylight bring about a substantial change in the distribution of light reflected from the surface of an object. (From Evans 1948.)
Figure 29-8 The reflectance function of a natural surface has component functions. A Three component functions, when added together in suitable proportions, provide the best three variable description of the surface reflectance functions of a large sample of natural objects. One curve can be loosely considered to represent the brightness dimension of the image; two other curves can be loosely thought of as representing dimensions of red-green variation and yellow-blue variation. These three component functions account for over 99% of the variance in the reflectance functions of the natural surfaces that have been studied. The fit can be made almost perfect by using three additional component functions (making six altogether). (From Cohen 1964 and Maloney 1986.) B The surface reflectance of an apple was measured at the point marked by the cross. C The apple's actual surface reflectance function is closely approximated by the best-fitting curve (dashed line) that could be synthesized by adding together, in appropriate proportions, the three fixed curves from A. The reflectance functions of most natural surfaces vary relatively smoothly and slowly with wavelength (see Figure 29-4A, B). The curve that describes surface reflectance can actually be assembled from a small number of even simpler underlying curves that can be thought of as elementary constituents common to all reflectance functions. Figure 29-8 shows a set of three such curves. By adding these three fundamental curves together in appropriate proportions, we can synthesize a real surface reflectance function rather well; by adding yet more fundamental curves we could do better still, though each additional curve would contribute progressively less information about the structure of the full reflectance function. Thus a visual system with a few sensing mechanisms (perhaps two or three) and spectral sensitivities that allowed the system to represent the underlying curves faithfully could do a good job of representing the range of spectral reflectance functions found among natural surfaces. Lawrence Maloney has shown that three mechanisms with the spectral sensitivities of human cones can in fact do the job well. The spectral sensing capabilities of the human eye therefore seem to be reasonably well-matched to the demands of distinguishing natural surfaces.
Figure 29-10 Spectral sensitivities of three “second-stage” mechanisms that can transform signals from the three classes of cones. A. The spectral sensitivity of the achromatic mechanism ( left ) is formed by adding signals from L and M cones ( right ), and possibly a very small contribution from S cones. The spectral sensitivity curves for the components ( right ) are the same as those shown in Figure 29-7 but are drawn on a linear rather than logarithmic sensitivity axis. B The spectral sensitivity of the red-green mechanism results from the subtraction of M cone signals from L cone signals. Some psychophysical observations suggest that S cones contribute to this mechanism, with the same sign as L cones, although this has not been found in studies of neurons in the lateral geniculate nucleus. C The spectral sensitivity of the yellow-blue mechanism results from the subtraction of L cone and M cone signals from S cone signals. Although the human visual system will not generally confuse light distributions reflected from different natural surfaces, a three-valued representation leaves the system open to confusion by artificial means. We exploit this in color reproduction systems to render a range of colors with only a small number of primary sources. For example, in color television a wide range of colors and lightnesses is synthesized at each point in the image by varying the intensities that excite three phosphors on the surface of the tube: one phosphor that emits long-wavelength light, one that emits middle-wavelength light, and one that emits shortwavelength light (Figure 29-9). Signals From Cones Are Transformed Early in the Visual Pathway Although the three kinds of cones can capture and represent the reflectances of natural surfaces, information would not be conveyed efficiently in a system in which each type of cone was connected to its own distinct neural pathway. This is easy to see if we consider the signals that arise in the L and M cones exposed to the kinds of spectral distributions that characterize the samples of flowers and skin in Figure 29-4. Because the reflectance functions vary slowly across the spectrum, and the spectral sensitivities of the L and M cones are similar over a broad spectral region, these classes of cones will generate highly correlated signals when they absorb light from natural surfaces. The correlation between these signals and the signals from the S cones is lower, but still substantial. The visual pathway could transmit information more efficiently by first removing from the cone signals those parts that are shared. The simplest way to do this is to transmit the differences among signals. The best transformation would be one that yielded, for the kinds of visual stimuli the eye normally encounters, the smallest correlation among signals in three pathways. Given the spectral composition of the light entering the eye, and the spectral sensitivities of the cone photoreceptors, the best transformations are the following: The sum of the signals from the three classes of cones (L+M+S). ● The difference between the signals from L and M cones (L - M). ● The difference between the signal from S cones and some combined signal from the L and M cones (S - LM). Mechanisms that transformed cone signals in these ways would have spectral sensitivities broadly like those shown in Figure 29-10. This result is very important, for it provides evidence for a transformation of cone signals that had long been suspected from perceptual observations and that has more recently been explored in physiological experiments.
Transmission of Color Signals by the Ganglion Cells A single ganglion cell may be stimulated by several cones or by only a few. When all three types of cones— the red, blue, and green types—stimulate the same ganglion cell, the signal transmitted through the ganglion cell is the same for any color of the spectrum. Therefore, the signal from the ganglion cell plays no role in the detection of different colors. Instead, it is a “white” signal. Conversely, some of the ganglion cells are excited by only one color type of cone but inhibited by a second type. For instance, this frequently occurs for the red and green cones, with red causing excitation and green causing inhibition, or vice versa. The same type of reciprocal effect occurs between blue cones on the one hand and a combination of red and green cones (both of which are excited by yellow) on the other hand, giving a reciprocal excitation-inhibition relation between the blue and yellow colors. The mechanism of this opposing effect of colors is the following: One color type of cone excites the ganglion cell by the direct excitatory route through a depolarizing bipolar cell, whereas the other color type inhibits the ganglion cell by the indirect inhibitory route through a hyperpolarizing bipolar cell. The importance of these color-contrast mechanisms is that they represent a means by which the retina itself begins to differentiate colors. Thus, each color-contrast type of ganglion cell is excited by one color but inhibited by the “opponent” color. Therefore, color analysis begins in the retina and is not entirely a function of the brain. Figure 29-11 The receptive fields of primate retinal ganglion cells have two concentrically organized regions, a center and an antagonistic surround. This fundamental organization is expressed in two basic forms: In an on-center cell, light falling on the center excites the cell while light falling on the surround inhibits it. In an off-center cell, light falling on the center inhibits the cell while light falling on the surround excites it. Different types of ganglion cells are distinguished by the sizes of their receptive fields and by the ways in which their centers and surrounds integrate signals from the different classes of cones. A M cells constitute about 8% of all ganglion cells. Even the smallest center receives inputs from several cones. The spectral sensitivities of center and surround differ little, if at all. B P cells constitute about 80% of all ganglion cells. Two subtypes are defined by the organization of cone inputs. The “red-green” opponent type receives inputs only from L and M cones, whereas the “yellow-blue” type receives input from all three classes of cones. Within the red-green class different connections to cones give rise to multiple cell subtypes: on- or off-center cells, and centers with L or M cone inputs. The cone inputs to the surround are less firmly established, but are generally thought to arise from the class that does not feed the center. In cells in and near the fovea the center of the receptive field receives input from a single cone; in more peripheral regions of retina several cones provide input to the center. Less is known about the structure of the receptive fields of the blue-yellow type of P cell. The antagonistic mechanisms seem to overlap more, possibly because chromatic aberration defocuses short-wavelength light, and center and surround are harder to distinguish. Cells in which S cones contribute the “off” signal are rare. In the late nineteenth century Ewald Hering first drew attention to the fact that the hues red, yellow, green, and blue have special properties: They are fundamental in the sense that other hues can readily be described as mixtures of them, and they seem to be related in mutually exclusive pairs (red vs green and blue vs yellow), so that a reddish-green color for example is impossible. These observations led Hering to postulate that vision depended on three distinct opponent mechanisms. One captured red-green variation in the image, so that it might be excited by red light and inhibited by green light (or vice-versa); another captured blue-yellow variation in the image, being perhaps excited by blue light and inhibited by yellow light. A third captured the light-dark, or achromatic , variation in the image, being excited by light and inhibited by dark. For a considerable time this account was seen as an alternative to the one that postulated three sensing systems of the kind represented by cones, but it eventually came to be seen as a description of mechanisms that receive and transform cone signals. It gained widespread acceptance in the 1950s when Leo Hurvich and Dorothea Jameson marshaled substantial evidence from psychophysical experiments and several other investigators found direct physiological evidence for the existence of mechanisms that combined cone signals in different ways. The first electrophysiological evidence for opponent mechanisms came from recordings made from horizontal cells in the fish retina. These cells become hyperpolarized in response to lights of certain wavelengths and are depolarized in response to lights of other wavelengths. Recordings in the primate retina by Dennis Dacey and colleagues show that the horizontal cells behave differently, giving responses of the same polarity to lights of all wavelengths. Opponent inputs are clearly evident in extracellular recordings made from primate ganglion cells by Peter Gouras and from lateral geniculate nucleus cells by Russell DeValois and colleagues and by Wiesel and Hubel. The properties of neurons in the lateral geniculate nucleus apparently reflect those of ganglion cells, which in turn probably reflect transformations of cone signals occurring at an earlier stage in the retina. Anatomical considerations suggest these happen in bipolar cells, though this has not yet been established physiologically. How do ganglion cells convey the red-green, blue-yellow, and achromatic dimensions of variation in the image? There are several kinds of retinal ganglion cells that differ in their anatomical and physiological characteristics. As we have seen in Chapter 26, there are two major classes of ganglion cells, now most often called M cells and P cells for their separate projections to the magnocellular (large-cell) and parvocellular (small-cell) layers of the lateral geniculate nucleus. These account for about 90% of all ganglion cells. Table 29-1 summarizes some of their properties, and Figure 29-11 shows schematically the organization of receptive fields. Modern physiological recordings from ganglion cells, and from the neurons to which they project in the lateral geniculate nucleus, have firmly established the chromatic properties of both M and P cells. The receptive fields of M cells have a simple antagonistic center-surround organization. Some cells have on-center receptive fields, others off-center receptive fields, but in both types the center and surround have similar, broad spectral sensitivities. Information about color is carried almost exclusively in the P cell system. P cells fall into two subtypes: neurons that receive opposed signals from L and M cones and neurons that receive signals from S cones opposed to some combined signal from L and M cones. These two subtypes of P cells are well suited to provide the red-green and blueyellow channels postulated by Hering, while M cells appear well equipped to convey the achromatic/ brightness signals. However, this scheme looks less satisfactory when we consider how the different kinds of neurons deal with the spatial structure of the image. Most of the information about the detailed structure of the visual world is conveyed by variations of brightness in the image rather than by variations in color (see Figure 29-1). We might therefore expect much of the visual system's capacity to be devoted to analyzing the brightness variations rather than the color variations. Perceptual experiments confirm this expectation: our capacity to resolve fine spatial variations in brightness exceeds our capacity to resolve spatial variations in hue. Since we know how M cells and P cells are distributed on the retina, we can calculate what kind of image detail each class of cell is capable of conveying. M cells are arranged much too sparsely to account for our capacity to resolve detail. They are important in the analysis of image movement (see Chapter 28), but they cannot convey information about detailed spatial variations in brightness. P cells, on the other hand, are very densely distributed—in and around the fovea there are two P cells for every cone— and could easily represent the detail that we can resolve in an image. The problem is that P cells appear superficially to be better suited to conveying information about color than to conveying information about the lightness variations that define structure. In fact, however, the spatial organization of the P cell's receptive field allows the cell to convey both brightness and color information in a complex signal. When the light falling on a P cell's receptive field covers both center and surround, the cell will respond well to variations in color, being excited by some hues and inhibited by others. However, when the light is either very small and confined to the center or distributed over the whole receptive field in a way that does not disturb the average light level on the surround (for example a fine grating pattern), only the central part of the receptive field generates a signal. Because this signal arises from a single type of cone, the cell responds to light over a broad range of wavelengths—it loses its color opponency. Thus a P cell responds well to brightness variations in the fine structure of the image, and it responds well to color variations in the coarse structure of the image. Not all P cells contribute to this encoding of spatial variations in lightness. The cornea and lens of the eye, when imaging a surface, cannot focus light of all wavelengths in the same plane in the retina. This chromatic aberration prevents all wavelengths from being in focus at the same time, so the visual system opts for a sharp image at middle and long wavelengths at the expense of a blurred image at short wavelengths. The S cones constitute less than 10% of the total (they are actually entirely absent from the center of the fovea). The ganglion cells that receive their inputs probably constitute a similar fraction of P cells and are sparsely distributed on the retina. P cells that receive inputs from only L and M cones seem to be the ones that can carry both a color-opponent signal and a brightness signal. These signals are conveyed through the ongoing discharge of action potentials and, as far as we know, are confounded in the discharge of a single cell. That is, just as individual cones confuse variations in wavelength and intensity, so too do individual P cells. The ambiguity in the discharge of any one P cell must be resolved by mechanisms in the cortex.
Signals Are Transformed Again in the Primary Visual Cortex Psychophysical observations suggest that information about color is encoded in the cortex in ways more complex than in the retina and lateral geniculate nucleus. The Cortex Contains More Than Three Chromatic Channels Although the idea that there are three “second-stage” mechanisms of color vision is now widely accepted, several lines of evidence suggest that this is a simplification and that, at least at higher levels in the visual pathway, many mechanisms exist, each selectively sensitive to its own small domain of color and lightness. The strongest evidence for this kind of organization comes from psychophysical experiments that show that sensitivity to particular hues is diminished after prolonged viewing of similar and complementary hues and that the loss of sensitivity is confined to spectral regions so narrow as to implicate more than three spectrally selective mechanisms. These experiments do not give a precise indication of how many higher-level mechanisms might exist, but they point firmly to more than three. Color is not an isolated attribute detached from other properties of an object, such as shape and movement. It is inextricably bound up with other object attributes, a point emphasized by perceptual tests that show how spatial context affects the appearance of colored regions (see Figure 29-3). When we consider the spatial complexities of natural scenes, the color of surfaces can no longer be treated simply as a three-variable problem. We know only a little about how the spatial attributes of objects influence their color, but it seems likely, given the simple behavior of P cells, that the mechanisms responsible for the influence of context reside in the cortex. Neurons in Primary Visual Cortex Do Not Fall Into Distinct Color Classes The P cells project from the lateral geniculate nucleus to neurons in layers 4Cβ and 4A of the primary visual cortex. Most of these neurons respond well to achromatic stimuli and poorly to colored ones. Some cells in layer 4Cβ, however, have receptive fields that resemble those of P cells: They have a concentrically organized color-opponent structure, though in some the antagonistic mechanisms are spatially coextensive. As a result, these cells in layer 4Cβ are most sensitive to changes in the color of a uniform region of light that covers the whole receptive field. They are relatively insensitive to brightness changes in either finely structured or coarsely structured stimuli. Layer 4A also contains a small proportion of cells with similarly organized color-opponent receptive fields. Thus, even in the input layers of the cortex there is substantial transformation of the signals arriving from the lateral geniculate nucleus, and there are probably cells (unlike those in the lateral geniculate nucleus) whose chromatic properties do not depend upon the spatial configuration of the visual stimulus. Neurons in the upper layers of striate cortex, to which layer 4 cells project heavily and which provide the principal output to higher visual areas, generally have receptive fields with a more elaborate structure. Most simple and complex cells (see Chapters 27 and 28) respond best to achromatic stimuli. Few cells respond well to color variations, even if these variations are configured to match the spatial characteristics of the cell's receptive field. We should not think this surprising, given the relatively small amount of information carried by color variation in images. Some of the cells that respond well to colored stimuli are simple, some are complex. Still other, rare ones have distinctive receptive fields consisting of concentrically organized regions. In the central region certain colors excite the cell while other colors inhibit it. Surrounding the central region is a much larger zone in which light of broad spectral composition will reduce the response (either excitatory or inhibitory) to light in the center. Neurons with fields like this are sometimes called double-opponent cells. Unlike the tight clustering of P cells in the lateral geniculate nucleus into clear “red-green” and “blue-yellow” groups, cells in the primary visual cortex that respond well to changes in color do not fall into distinct groups and are widely scattered, as if each cell is selective for a particular combination of brightness and color contrast. The code the brain uses to convey information about color is profoundly charged in the primary visual cortex. The general picture emerging from studies of striate cortex is that color is just one of several dimensions of image variation to which an individual neuron is selective; information about color is encoded along with information about other attributes of the image. However, this picture is complicated by the possibility that color-opponent cells are clustered in the striate cortex. Some investigators have found relatively high concentrations of double-opponent cells in the conspicuous “blobs” that regularly punctuate the striate cortex. Because blobs send specific projections to higher visual areas (Chapter 27), any clustering of color-opponent cells in blobs implies the existence of cortical pathways that analyze color. Signals About Color Are Conveyed to the Temporal Lobe The secondary visual area, V2, is the principal destination of output from the primary visual cortex. When stained for cytochrome oxidase, V2 in the monkey shows a pattern of stripes, and some of these (the “thin” stripes) receive inputs preferentially from the blobs in striate cortex (see Chapter 28). The thin stripes in turn send projections to area V4, a region that Semir Zeki first showed contained many cells that are selective for the color of visual stimuli. As a result of Zeki's observations, V4 is regarded as an area whose principal role is to analyze and represent the color information in the image. As discussed in Chapter 28, Zeki's discoveries, in both V4 and MT (the middle temporal area), indicate that different visual cortical areas are specialized for the analysis of different attributes of the image: color, motion, depth, etc. To the extent that color opponent neurons in V1 are associated with the blobs, this evidence points to a pathway specialized for the analysis of color. Thus, information about the chromatic attributes of objects is confined mainly to pathways that convey information from striate cortex, through areas V2 and V4, to the temporal lobe. Experimental studies of animals with lesions of these pathways, and studies of people who have suffered localized brain damage, usually as a result of stroke, sometimes show impairments of color vision, often in association with other disruptions of object vision. Recordings made from single neurons at different stages in the pathway connecting the primary visual cortex to the temporal lobe show that their chromatic characteristics are generally like those of neurons in the primary cortex. Some studies of neurons in area V4 have found that a cell's response to a colored stimulus falling on its receptive field is influenced by the color of light falling in a large region surrounding the receptive field. This work implicates V4 in color-contrast phenomena of the kind illustrated in Figure 29-3. The Importance of Context in Color Perception Seeing the luminance of objects (that is, their brightness) can presumably be signaled by simply increasing or decreasing the overall firing rate of the relevant retinal ganglion cells, properly adapted to the overall level of ambient light (see text and Box E ). Seeing color, however, logically demands that retinal responses to different wavelengths in some way be compared . The discovery of three human cone types and their different absorption spectra is correctly regarded, therefore, as the basis for human color vision. Nevertheless, how the three human cone types and the higher-order neurons they contact (see Chapter 12 ) produce the sensations of color is still unclear. Indeed, this issue has been debated by some of the greatest minds in science (Hering, Helmholtz, Maxwell, Schroedinger, Mach, and Land, to name only a few) since Thomas Young first proposed that humans must have three different receptive “particles,” i.e., cone types. A fundamental problem has been that, although the relative activities of three cone types can more or less explain the colors perceived in color matching experiments performed in the laboratory, the perception of color is strongly influenced by context. For example, a patch returning the exact same spectrum of wavelengths to the eye can appear quite different depending on its surround, a phenomenon called color contrast (see figure). Moreover, test patches returning different spectra to the eye can appear to be the same color, an effect called color constancy . Although these phenomena were well known in the nineteenth century, they were not accorded a central place in color vision theory until Edwin Land's work in the 1950s. In his most famous demonstration, Land (who among other achievements founded the Polaroid company and became a billionaire) used a collage of colored papers that have been referred to as “the Land Mondrians” because of their similarity to the work of the Dutch artist Piet Mondrian. Using a telemetric photometer and three adjustable illuminators generating short, middle, and long wavelength light, Land showed that two patches that in white light appeared quite different in color (e. g., green and brown) continued to look their respective colors even when the three illuminators were adjusted so that the light being returned from the “green” surfaces produced exactly the same readings on the three telephotometers as had previously come from the “brown” surface—a striking demonstration of color constancy! The phenomena of color contrast and color constancy have led to a heated debate about how color percepts are generated that now spans several decades. For Land, the answer lay in a series of ratiometric equations that could integrate the spectral returns of different regions over the entire scene. It was recognized even before Land's death in 1991, however, that his so-called retinex theory did not work in all circumstances and was in any event a description rather than an explanation. An alternative explanation of these contextual aspects of color vision is that color, like brightness, is generated empirically according to what spectral stimuli have typically signified (see Box E ). The brown tile at the center of the illuminated upper face of the cube and the orange tile at the center of the shadowed face are actually returning the same spectral light to the eye (as is the tan tile lying on the ground-plane in the foreground). Readers who find this hard to believe can convince themselves by cutting holes in a sheet of paper such that the rest of the scene is masked out, in which case the two tiles on the faces of the cube look identical in both color and brightness. This illustration provides a dramatic example of the influence of context on the color perceived. (From Lotto and Purves, 1999.)
Color Blindness Red-Green Color Blindness. When a single group of colorreceptive cones is missing from the eye, the person is unable to distinguish some colors from others. For instance, one can see in Figure 50–9 that green, yellow, orange, and red colors, which are the colors between the wavelengths of 525 and 675 nanometers, are normally distinguished from one another by the red and green cones. If either of these two cones is missing, the person cannot use this mechanism for distinguishing these four colors; the person is especially unable to distinguish red from green and is therefore said to have red-green color blindness. A person with loss of red cones is called a protanope; the overall visual spectrum is noticeably shortened at the long wavelength end because of a lack of the red cones. A color-blind person who lacks green cones is called a deuteranope; this person has a perfectly normal visual spectral width because red cones are available to detect the long wavelength red color. Red-green color blindness is a genetic disorder that occurs almost exclusively in males. That is, genes in the female X chromosome code for the respective cones. Yet color blindness almost never occurs in females because at least one of the two X chromosomes almost always has a normal gene for each type of cone. Because the male has only one X chromosome, a missing gene can lead to color blindness. Because the X chromosome in the male is always inherited from the mother, never from the father, color blindness is passed from mother to son, and the mother is said to be a color blindness carrier; this is true in about 8 per cent of all women. Blue Weakness. Only rarely are blue cones missing, although sometimes they are underrepresented, which is a genetically inherited state giving rise to the phenomenon called blue weakness. Color Test Charts. A rapid method for determining color blindness is based on the use of spot charts such as those shown in Figure 50–10. These charts are arranged with a confusion of spots of several different colors. In the top chart, the person with normal color vision reads “74,” whereas the red-green color-blind person reads “21.” In the bottom chart, the person with normal color vision reads “42,” whereas the red-blind person reads “2,” and the green-blind person reads “4.” If one studies these charts while at the same time observing the spectral sensitivity curves of the different cones depicted in Figure 50–9, it can be readily understood how excessive emphasis can be placed on spots of certain colors by color-blind people.